
Белокровные рыбы
- 1 year ago
- 0
- 0

Носо́рогие белокро́вки , или кергеленские носорогие белокровки ( лат. Channichthys ) — род морских субантарктических донных и придонных рыб семейства белокровковых (Channichthyidae) отряда окунеобразных (Perciformes). Латинское название рода, данное ему в 1844 году шотландским натуралистом и ихтиологом Джоном Ричардсоном , происходит от двух латинизированных греческих слов — «анчоус» ( channe ) и «рыба» ( ichthys ). В состав рода включают 9 видов .
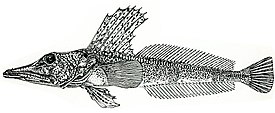
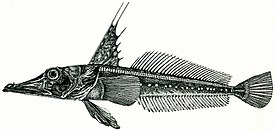
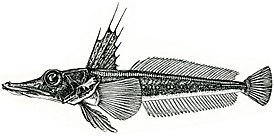
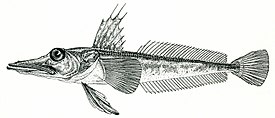
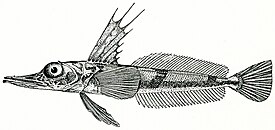
Русское название «носорогие белокровки» и английское название «unicorn fishes» отражают наличие у этих рыб хорошо развитого шипа («рога»), венчающего передний край рыла. Иногда носорогих белокровок называют «носорожьими белокровками» — неудачно переведенным с английского названием. Подобный перевод некорректен с этимологической точки зрения, так как обозначает принадлежность какого-либо объекта к носорогу или внешнюю схожесть с ним, тогда как основной смысл названия должен отражать одну из самых характерных черт внешнего вида этих рыб — наличие на кончике рыла хорошо развитого шипа.
Характерной особенностью носорогих белокровок, кроме наличия рострального шипа, является, как и у всех остальных видов семейства — отсутствие на теле чешуи, за исключением плотных костных чешуй в боковых линиях, и наличие бесцветной крови, лишенной зрелых форм эритроцитов и гемоглобина — уникальное явление среди всех позвоночных, присущее только 25 видам рыб этого семейства. Кровь белокровковых рыб, по сути, представляет собой слегка желтоватую плазму. Редукция эритроцитов, а также гемоглобина и мышечного пигмента миоглобина у белокровковых рыб объясняется эволюционными изменениями, происходившими в организме предковых форм на фоне относительно резкого похолодания климата в южнополярном регионе после отделения Антарктиды от Южной Америки и соответственного снижения температуры воды в Южном океане до отрицательных значений (до точки −1,9 °C). С физиологической точки зрения это явление объясняется уменьшением плотности крови за счет редукции её форменных элементов, что заметно уменьшает плотность, а также снижает вязкость жидкости при низких температурах и соответственно повышает её текучесть в сосудах кровеносной системы .
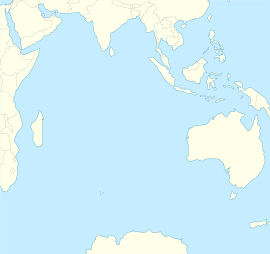
Носорогие белокровки — среднего и крупного размера рыбы (общей длиной до 61 см) являются эндемиками антарктических морских вод у северной границы Южного океана , омывающих острова архипелага Кергелен и Хёрд , а также серии подводных поднятий — гайотов (банок) между ними, расположенными в Индоокеанском секторе Антарктики в районе подводного хребта Кергелен .
Встречаются в качестве прилова при промысле в районе островов Кергелен щуковидной белокровки Chamsocephalus gunnari Lönnberg, 1905, больше известной под коммерческим названием « ледяная рыба ».

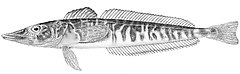
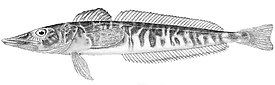
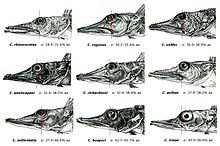
Ростральный шип хорошо развит, его вершина направлена назад. Верхняя поверхность головы, а у некоторых видов и лучи первого спинного и брюшного плавников, обычно покрыты мелкими костными гранулами — туберкулами, придающими поверхности шершавость; у некоторых видов костные туберкулы встречаются на всем теле. Крышечная кость с 5—7 мощными шипами. Две боковых линии — дорсальная (верхняя) и медиальная (срединная), более короткая, в которых присутствуют трубчатые или прободенные плотные костные членики ( чешуи ) . Два спинных плавника широко разделены междорсальным промежутком либо сближены вплотную. Первый спинной плавник высокий или очень высокий, содержит 5—11 гибких колючек; во втором спинном плавнике 30—36 членистых лучей, в анальном плавнике 28-33 членистых лучей, в грудном плавнике 18—23 луча. Третий луч брюшного плавника наиболее длинный. Хвостовой плавник закругленный. Лучей жаберной перепонки 6. Тычинки на первой жаберной дуге рудиментарные или хорошо развиты, покрыты мелкими зубчиками и могут быть расположены в 1 или 2 ряда — внешнем и внутреннем; общее число тычинок широко варьирует у разных видов от 6 до 31 .
Ареал рода охватывает острова и подводные поднятия подводного хребта Кергелен от северной его оконечности — островов Кергелен, до южной — островов Хёрд и Макдональдс. Согласно схеме зоогеографического районирования по донным рыбам, предложенной А. П. Андрияшевым и А. В. Нееловым , виды носорогих белокровок являются эндемиками округа Кергелен-Хёрд Индоокеанской провинции Антарктической области. Все 9 видов известны и описаны от островов Кергелен, а парусная белокровка C. velifer Meissner, 1974 была также обнаружена на изолированных поднятиях (банках) подводного хребта Кергелен. Сведения о точной идентификации рыб из остальной части ареала рода отсутствуют. В районе островов Хёрд и Макдональдс, в южной части ареала, представители этого рода традиционно упоминаются в литературе под сборным названием C. rhinoceratus Richardson, 1844 .
Относительно мелководные виды, известные глубин 95—361 м по результатам донных тралений у островов Кергелен и на поднятиях подводного хребта Кергелен.
Наиболее крупными видами рода являются C. rhinoceratus , достигающая общей длины 61 см, и C. velifer с общей длиной до 54 см. В среднеразмерную группу входят C. mithridatis , достигающая общей длины 44 см, C. bospori , достигающая общей длины не менее 41 см, C. panticapaei с общей длиной до 40 см, C. richardsoni — не более 38 см и C. aelitae — до 37 см. Самые мелкие виды — C. irinae , общая длина которой не превышает 26 см, и C. rugosus с общей длиной до 31 см .
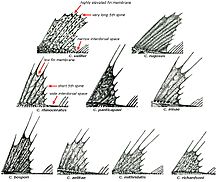
По приуроченности носорогих белокровок к определенным биотопам , согласно особенностям их внешнего строения и особенностям питания, среди них выделяются 2 группы рыб — типично донные хищники — и ихтиофаги , вероятно, проводящие большую часть жизни на поверхности грунта ( белокровка Ричардсона C. richardsoni Shandikov, 2012 , рыжая белокровка C. rugosus Regan, 1913 и C. velifer ), и придонные или придонно-пелагические — хищники -ихтиофаги и зоопланктофаги , имеющие более пелагизированную форму тела, которые для питания, очевидно, периодически поднимаются в толщу воды ( большеглазая белокровка C. bospori Shandikov, 1995 , карликовая белокровка C. irinae Shandikov, 1995 , зелёная белокровка C. mithridatis Shandikov, 2009 , угольная белокровка C. panticapaei Shandikov, 1995 , носорогая белокровка C. rhinoceratus и, вероятно, белокровка Аэлиты C. aelitae Shandikov, 1995 ) .
В питании донных видов C. rugosus и C. velifer отмечена рыба и головоногие моллюски. В питании придонных и придонно-пелагических видов наблюдается четкое разделение на 2 группы видов — хищников и зоопланктофагов. К хищникам, специализирующимся на ихтиофагии, относятся C. rhinoceratus , C. mithridatis , C. velifer и C. aelitae , а к типичным зоопланктофагам, питающимся амфиподами ( Themisto gaudishaudi ) и мелкими эвфаузиевыми ( Thysanoessa macrura ), относятся C. irinae , C. bospori и C. panticapaei . Хищным рыбам свойственно более низкое число (6-16) относительно слаборазвитых жаберных тычинок, расположенных только во внешнем ряду первой жаберной дуги. Тогда как у зоопланктофагов тычинки более развиты и плотнее покрыты костными шипиками, их большее количество (18-31) и расположены они в 2 ряда — внешнем и внутреннем.
Размеры впервые созревающих рыб, сезоны и сроки нереста заметно различаются у разных видов. Поэтому близкие к нересту рыбы рода Channichthys встречаются в водах, омывающих острова Кергелен, в течение круглого года. Преднерестовые самки C.rhinoceratus впервые созревают при общей длине около 49 см, нерестится вид, очевидно, в конце лета южного полушария — в феврале-марте. C. velifer становится половозрелой, по-видимому, при общей длине около 31-33 см, размножается в летний период — в январе-феврале. У C. panticapaei половозрелость наступает при общей длине около 30 см; нерест, очевидно, приходится на зимний период — июнь-июль. C. bospori очевидно нерестится в июне-июле, размеры рыб при наступлении половозрелости неизвестны. У C. aelitae нерест происходит в летний период, скорее всего в январе-феврале, размеры впервые созревающих рыб не известны. Половозрелость у C. mithridatis наступает при общей длине 30-32 см, нерестится в зимний период, предположительно в мае-июне. C. richardsoni впервые созревает при общей длине около 29-31 см, нерест, очевидно, происходит в осенне-зимний период. C. rugosus впервые начинает созревать при общей длине не менее 28 см; нерест, вероятно, приходится на осенние месяцы. C. irinae впервые созревает при достижении общей длины 23-25 см; нерест происходит в зимний период — в июле-августе.
Абсолютная плодовитость, известная по одной самке C. velifer , составляет 7155 икринок.
В роде в настоящее время насчитывается 9 видов:
|
|
|
|---|