
Эпигенетика
- 1 year ago
- 0
- 0

Парамута́ция ( англ. paramutation ) — взаимодействие двух аллелей одного локуса , при котором один аллель вызывает наследуемые изменения в другом аллеле . Эти изменения могут заключаться в изменении паттерна метилирования ДНК или модификации гистонов . Аллель, индуцирующий эти изменения, называется парамутагенным, а тот аллель, который эпигенетически изменяется, называется парамутабельным . Парамутабельный аллель может иметь изменённые уровни экспрессии , которые могут сохраняться у потомства, унаследовавшего этот аллель, даже в отсутствие парамутагенного аллеля . Парамутации могут иметь место, например, у генетически идентичных растений , демонстрирующих совершенно разные фенотипы .
Хотя явление парамутации было изучено в основном на кукурузе , парамутации были описаны и у других организмов, включая таких животных , как плодовая мушка Drosophila melanogaster и мышь . Несмотря на широкое распространение, примеры парамутаций немногочисленны, и их механизмы не до конца понятны.
Первое описание явления, которое можно было бы назвать парамутацией, было сделано Уильямом Бэтсоном и Каролин Пеллью ( англ. Caroline Pellew ) в 1915 году. Они описали «карликовый» ( англ. rogue ) горох, который передавал свою «карликовость» потомству . Однако первое формальное описание парамутации было выполнено ( англ. R. A. Brink ) из Висконсинского университета в Мадисоне в 1950 году у растений кукурузы . Он заметил, что особые слабо экспрессируемые аллели локуса red1 ( r1 ) у кукурузы, который кодирует транскрипционный фактор , который придаёт красный цвет зёрнам кукурузы, может вызывать наследуемые изменения в сильно экспрессируемых аллелях, которые превращают их в слабо экспрессируемые. Слабый уровень экспрессии сохраняется при передаче этих аллелей потомству, и они могут, в свою очередь, понижать уровень экспрессии других аллелей (это явление получило название вторичной парамутации). Бринк показал, что влияние парамутагенного аллеля может передаваться нескольким последующим поколениям .
Аллели, которые могут вызывать наследуемые изменения в других аллелях, называются парамутагенными, а изменяющеся аллели называются парамутабельными. Аллели, не принимающие участия в этих взаимодействиях, называют нейтральными. Когда парамутагенный и парамутабельный аллели имеются у одного и того же организма, парамутабельный аллель превращается в парамутагенный и сохраняет свою парамутагенность в нескольких поколениях. При описанной трансформации не происходит никаких изменений в ДНК , однако парамутагенные и парамутабельные аллели различаются по эпигенетическим признакам (в том числе по метилированию ДНК). Как правило, парамутабельный аллель активно транскрибируется , а парамутагенный аллель транскрибируется мало или не транскрибируется вовсе. Первым описанным и наиболее изученным примером парамутации является локус r1 кукурузы. Ген , расположенный в этом локусе, когда транскрибируется, даёт начало транскрипционному фактору, стимулирующему образование антоциана , в результате чего зёрна приобретают пурпурную окраску. Один аллель этого локуса, обозначаемый B', может вызывать метилирование другого аллеля, B-I. Метилирование снижает транскрипцию и, соответственно, уровень синтеза антоциана. Эти аллели не отличаются по последовательностям ДНК, однако различаются по степени метилирования ДНК. Как и в случае других парамутаций, превращение аллели B-I в аллель B' стабильно и наследуемо. Другие похожие примеры парамутаций имеют место в случае некоторых других локусов кукурузы, а также у модельного растения Arabidopsis thaliana и трансгенных петуний . Парамутации также описаны у Drosophila melanogaster , червя Caenorhabditis elegans и мыши .
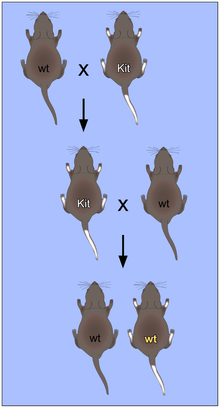
В качестве примера парамутаций можно рассмотреть парамутацию Kit у мышей . Ген Kit кодирует , который участвует в таких процессах развития, как гемопоэз , дифференцировка клеток зародышевой линии и меланогенез . Гомозиготная делеция Kit летальна, а гетерозиготные мыши имеют белый хвост. Белый хвост гетерозиготных родителей сохраняется у их потомков, имеющих генотип дикого типа (Kit +/+ ). Показано, что в этом случае парамутация обусловлена РНК : инъекция в оплодотворённое яйцо с генотипом дикого типа РНК из спермы белохвостой гомозиготы или РНК, образованной из транскрипта Kit , приводит к развитию белохвостого фенотипа .
Хотя специфические механизмы осуществления парамутаций различаются у разных организмов, в их основе лежат эпигенетические модификации и РНК-сайленсинг .
В случае локуса r1 кукурузы для парамутагенного аллеля B' характерно ДНК-метилирование участка тандемных повторов рядом с кодирующей областью, и когда парамутабельный аллель B-I становится парамутагенным, он тоже приобретает характерный паттерн ДНК-метилирования . Для успешного переноса ДНК-метилирования необходим ряд генов, кодирующих и другие белки пути РНК-сайленсинга, подтверждая тем самым, что парамутация осуществляется посредством эндогенного . Эта гипотеза подтверждается наличием транскрипции малых интерферирующих РНК с участка тандемных повторов. У таких животных, как D. melanogaster , парамутации могут быть опосредованы piРНК . Также было показано, что в некоторых случаях помимо ДНК-метилирования для осуществления парамутаций необходимы модификации гистонов метилированной ДНК, поэтому для парамутаций могут быть необходимы гистон-модифицирущие белки . Было высказано предположение, что модификации гистонов могут быть необходимы для поддержания парамутаций . Однако стоит отметить, что с помощью концепции РНК-сайленсинга невозможно объяснить все свойства парамутаций, поэтому в парамутациях могут быть задействованы и другие механизмы .
Существует предположение, что в отдельной популяции относительно немного генов демонстрируют парамутации ввиду того, что высокая пенетрантность парамутагенных аллелей может привести к закреплению или парамутагенного, или парамутабельного аллелей. Впрочем, парамутации могут сохраняться парамутации в локусах с низкой пенетрантностью парамутагенного аллеля. Это необходимо учитывать селекционерам растений .
Поскольку парамутации и явления, похожие на них, описаны у таких животных, как дрозофила и мышь, предполагается, что парамутации могут вносить свой вклад в развитие наследственных заболеваний человека с неменделевским наследованием .






