
Джейн, Кори
- 1 year ago
- 0
- 0
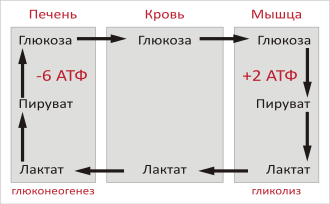
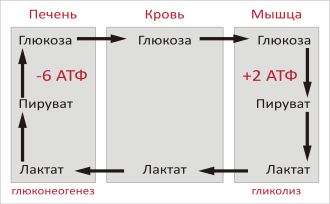

Цикл Кори (также известный как цикл молочной кислоты или глюкозо-лактатный цикл ), названный в честь его первооткрывателей Карла Фердинанда Кори и Герти Кори , представляет собой метаболический путь, при котором лактат , вырабатываемый в результате анаэробного гликолиза в мышцах, транспортируется в печень и превращается в глюкозу , которая затем возвращается в мышцы и циклически метаболизируется обратно в лактат . Расширенное описание включает метаболические пути глюконеогенеза , глутаминовую кислоту (Glu), части цитратного цикла и цикл мочевины .
Скелетная мышца не способна снова превращать лактат в глюкозу даже в аэробных условиях: ей не хватает ферментов глюконеогенеза . По этой причине происходит циркуляция метаболитов между мышцами и печенью — последняя имеет соответствующий ферментный репертуар. В своем первоначальном виде этот круговорот органов назывался «циклом Кори». Расширенная форма того же самого, «глюкозо-аланиновый цикл», возможно, имеет большее значение, поскольку она одновременно предотвращает отравление мышц аммиаком , доставляя его в аппарат детоксикации печени (цикл мочевины).
При интенсивной мышечной работе, а также в условиях отсутствия или недостаточного числа митохондрий (например, в эритроцитах или мышцах ) глюкоза вступает на путь анаэробного гликолиза с образованием лактата . Лактат не может далее окисляться , он накапливается (при его накоплении в мышцах раздражаются чувствительные нервные окончания, что вызывает характерное жжение в мышцах). С током крови лактат поступает в печень . Печень является основным местом скопления ферментов глюконеогенеза (синтез глюкозы из неуглеводных соединений), и лактат идет на синтез глюкозы.
Реакция превращения лактата в пируват катализируется лактатдегидрогеназой, далее пируват подвергается окислительному декарбоксилированию или может подвергаться брожению .
В целом, на этапах гликолиза цикла образуется 2 молекулы АТФ за счет 6 молекул АТФ, расходуемых на этапах глюконеогенеза. Каждая итерация цикла должна поддерживаться чистым потреблением 4 молекул АТФ. В результате цикл не может продолжаться бесконечно. Интенсивное потребление молекул АТФ в цикле Кори переносит метаболическую нагрузку с мышц на печень.
Цикл Кори получил название по первооткрывателю — его открыла чешская ученая, лауреат Нобелевской премии Тереза Кори .
Важность цикла основана на предотвращении лактоацидоза во время анаэробных условий в мышцах. Однако обычно, прежде чем это произойдет, молочная кислота выводится из мышц в печень .
Кроме того, этот цикл важен для производства АТФ, источника энергии, во время мышечной нагрузки. Окончание мышечной нагрузки позволяет циклу Кори функционировать более эффективно. Это погашает кислородный долг, поэтому и цепь переноса электронов, и цикл лимонной кислоты могут производить энергию с оптимальной эффективностью .
Цикл Кори является гораздо более важным источником субстрата для глюконеогенеза , чем пища . Вклад лактата цикла Кори в общее производство глюкозы увеличивается с увеличением продолжительности голодания до наступления плато . В частности, после 12, 20 и 40 часов голодания у добровольцев-людей на глюконеогенез приходится 41 %, 71 % и 92 % производства глюкозы, но вклад лактата из цикла Кори в глюконеогенез составляет 18 %, 35 % и 36 % соответственно . Оставшаяся глюкоза вырабатывается в результате расщепления белка , мышечного гликогена , и глицерина в результате липолиза .
Препарат метформин может вызывать лактоацидоз у пациентов с почечной недостаточностью , поскольку метформин ингибирует печеночный глюконеогенез цикла Кори, в частности комплекса 1 дыхательной цепи митохондрий . Накопление лактата и его субстратов для производства лактата, пирувата и аланина, приводит к избытку лактата . Обычно избыток кислоты, который является результатом ингибирования комплекса митохондриальных цепей, выводится почками, но у пациентов с почечной недостаточностью почки не могут справиться с избытком кислоты. Распространенное заблуждение гласит, что лактат является агентом, ответственным за ацидоз, но лактат представляет собой конъюгатное основание, которое в основном ионизируется при физиологическом рН и служит маркером образования кислоты, а не её причиной .
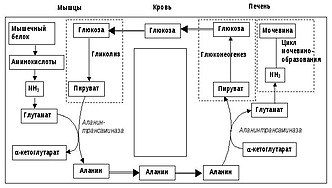
Белки расщепляются до аминокислот в цитозоле . Аминокислоты , в свою очередь, дезаминируются путем трансаминирования и помещают оставшиеся углеродные каркасы в цитратный цикл . Аминогруппа аминокислот временно переносится в кофактор пиридоксальфосфат (ПЛФ) при трансаминировании; Таким образом, ПЛФ превращается в пиридоксаминфосфат (ПАМФ). Аланинаминотрансфераза (АЛАТ, АЛТ) (также называемая глутамат-пируваттрансаминазой, ГПТ) переносит аминогруппировку ПАМФ на пируват в мышцах. Таким образом, образуется аланин и регенерированный ПЛФ, который, таким образом, может поглощать новые аминогруппы. Аланин транспортируется через кровь в печень, где АЛАТ из ПЛФ и аланина делает ПАМФ и пируват, который можно использовать для глюконеогенеза и отправлять обратно во внепеченочные клетки в виде глюкозы.
Через АЛАТ аминогруппа переносится из ПАМФ в α-кетоглутарат . Образующийся глутамат превращается в митохондриях клетки печени в α-кетоглутарат и NH 3 с помощью глутаматдегидрогеназы (ГЛДГ), последний превращается из с CO 2 в , который поступает в цикл мочевины. Вторая группа мочевины NH 2 поставляется через продукт трансаминирования аспартата (Asp), который, в свою очередь, расщепляется до аргинина и фумарата . Из аргинина в конечном итоге отделяется мочевина . Из фумарата можно регенерировать в аспартат через малат и оксалацетат ( ). Мочевина выводится через почки.
В отличие от цикла Кори, цикл аланина не только регенерирует углеводы, но и выводит NH 3 из мышц. Однако для этого в синтезе мочевины в печени также необходимо потратить энергию на утилизацию NH 3 .
| Гликолиз |
|
||||||||
|---|---|---|---|---|---|---|---|---|---|
| Только глюконеогенез |
|
||||||||
| Регуляция | |||||||||












