Транспорт в Чехии
- 1 year ago
- 0
- 0
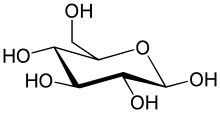
Глюкозные транспортёры ( англ. Glucose transporter , сокр. GLUT или ГЛЮТ ) — большая группа мембранных белков , отвечающих за перенос глюкозы через клеточную мембрану . Поскольку глюкоза является жизненно важным источником энергии, эти белки присутствуют у всех типов живых организмов.
Отдельно выделяют GLUT ( ГЛЮТ ), или — семейство белков-переносчиков глюкозы, встречающихся в большинстве клеток млекопитающих . Так, в человеческом геноме закодировано двенадцать белков семейства GLUT. Они представляют собой транспортные белки- унипортеры .
Большинство не автотрофных организмов неспособны производить свободную глюкозу, поскольку у них отсутствует экспрессия фермента глюкозо-6-фосфатазы . Таким образом, они способны осуществлять только поглощение и катаболизм глюкозы. Однако, в условиях голодания некоторые ткани и органы, такие как гепатоциты , кишечник , мышцы , мозг и почки , способны осуществлять синтез глюкозы, вследствие активации процесса глюконеогенеза .
В клетках модельного организма Saccharomyces cerevisiae транспорт глюкозы происходит путём . Большинство транспортных белков этого организма относятся к семейству Hxt, но есть и множество белков транспортёров из других семейств .
| Название | Характеристики | Описание |
| высокое сродство к глюкозе; подавляется глюкозой; низкий уровень экспресси; подавляет синтез Hxt6 | ||
| низкое сродство к глюкозе; низкий уровень экспресси | ||
| Km : 100 мM , 129 - 107 мM | низкое сродство к глюкозе; синтез индуцируется высоким уровнем глюкозы | |
| Km = 1.5 - 10 мM | высокое/среднее сродство к глюкозе; синтез индуцируется низким уровнем глюкозы | |
| Vm = 18.5, Kd = 0.078, Km = 28.6/34.2 - 60 мM | низкое сродство к глюкозе | |
| Vm = 12.0, Kd = 0.049, Km = 6.2 | среднее сродство к глюкозе | |
| Km = 10 мM | Среднее сродство к глюкозе. Сильная экспрессия в фазе стационарного роста, при образовании спор и в условиях низкой концентрации глюкозы. Транскрипция подавляется глюкозой . | |
| Vm = 11.4, Kd = 0.029, Km = 0.9/14 , 1.5 mM | высокое сродство к глюкозе | |
| Vm = 11.7, Kd = 0.039, Km = 1.3, 1.9, 1.5 mM | высокое сродство к глюкозе | |
| низкий уровень экспрессии | ||
| участвует в множественной лекарственной устойчивости | ||
| участвует в множественной лекарственной устойчивости | ||
| Vm = 17.5, Kd = 0.043, Km = 1.5, 1.6 | высокое сродство к галактозе |
GLUT ( ГЛЮТ ) — интегральные мембранные белки , содержащие 12 пересекающих клеточную мембрану спиралей, при этом амино- (N-конец) и карбоксильный (C-конец) концы выходят со стороны клеточной мембраны, обращенной к цитоплазме . ГЛЮТ переносят глюкозу и связанные гексозы в соответствии с моделью альтернативных конформаций , которая предсказывает, что транспортёр выставляет свой единственный участок связывания субстрата либо внутрь, либо наружу клетки. Связывание глюкозы с участком провоцирует конформационное изменение , связанное с транспортом, и приводит к высвобождению глюкозы с противоположной стороны клеточной мембраны. Внутренние и внешние участки связывания глюкозы, как считается, расположены на трансмембранных сегментах 9, 10 и 11 . QLS на седьмом трансмембранном сегменте возможно может определять селективность и аффинность транспорта .
Каждая изоформа глюкозного транспортёра играет определенную роль в метаболизме глюкозы, в зависимости от её тканевой экспрессии, субстратной специфичности, кинетики транспорта и регуляции экспрессии в различных физиологических условиях . На данный момент обнаружено тринадцать белков-транспортёров семейства GLUT/SLC2 . На основании сходства аминокислотных последовательностей они разделены на три подкласса.
К классу I относятся транспортёры GLUT1-GLUT4 .
| Название | Распространение | Описание |
| GLUT1 | Широко распространён в зародышевых тканях. У взрослых сильнее всего экспрессируется в эритроцитах и эндотелиальных клетках барьерных тканей, например в гемато-энцефалическом барьере . Помимо этого, он ответственен за минимальный базальный уровень поглощения глюкозы всеми клетками организма, необходимый для поддержания клеточного дыхания. | Уровень GLUT1 в клеточной мембране повышается при снижении уровня глюкозы и уменьшается при его повышении. |
| GLUT2 | Это транспортёр глюкозы, работающий в двух направлениях. Экспрессируется клетками почечных канальцев, печени и бета-клетками поджелудочной железы. Также его можно обнаружить в базолатеральной мембране эпителия тонкой кишки. Двунаправленный транспортёр необходим клеткам печени для поглощения глюкозы в процессе гликолиза , и её высвобождения в процессе глюконеогенеза . В бета-клетках поджелудочной железы, свободная глюкоза необходима для того, что бы клетки могли точно измерить уровень глюкозы в сыворотке крови. Кроме этого GLUT2 осуществляет транспорт глюкозы , галактозы и фруктозы из клеток слизистой кишечника в просвет кровеносных сосудов. | Это изоформа с низким сродством. Существуют данные, что основными транспортёрами глюкозы в бета-клетки на самом деле являются GLUT1 и GLUT3. |
| GLUT3 | В основном экспрессируется в нейронах (где, как полагают, он является главной изоформой глюкозного транспортёра) и плаценте . | Это изоформа с высоким сродством к глюкозе, что позволяет ей осуществлять транспорт при низких концентрациях глюкозы. |
| GLUT4 | Обнаружен в жировой ткани , а также в скелетных мышцах и миокарде . | Этот транспортёр регулируется инсулином . Осуществляет инсулин-зависимое поглощение глюкозы. |
К классу II относятся:
К классу III относятся:
Большинство транспортёров классов II/III было обнаружено относительно недавно в результате деятельности различных геномных проектов.
Функции данных изоформ на данный момент неясны. Некоторые из них (GLUT6, GLUT8) состоят из , которые способствуют сохранению транспортёров внутри клетки, и таким образом предотвращают транспорт глюкозы. Существуют ли механизмы, способствующие транслокации этих транспортёров на клеточную поверхность, неизвестно, но было выяснено, что инсулин не способствует такой транслокации.
В августе 1960 года, в Праге, представил общественности своё открытие: механизм вторично-активного транспорта глюкозы в сопряжении с натрием в клетках кишечника . Открытие Крэйном вторично-активного транспорта было первым открытием, показавшим значимость сопряжения потоков в биологии .
“ in 1961 was the first to formulate the cotransport concept to explain active transport [7]. Specifically, he proposed that the accumulation of glucose in the intestinal epithelium across the brush border membrane was [is] coupled to downhill Na+ transport cross the brush border. This hypothesis was rapidly tested, refined, and extended [to] encompass the active transport of a diverse range of molecules and ions into virtually every cell type.”
“the insight from this time that remains in all current text books is the notion of published originally as an appendix to a symposium paper published in 1960 ( et al. 1960). The key point here was 'flux coupling', the of sodium and glucose in the apical membrane of the small intestinal epithelial cell. Half a century later this idea has turned into one of the most studied of all transporter proteins (SGLT1), the sodium–glucose cotransporter.”