Региональное кодирование оптических дисков
- 1 year ago
- 0
- 0
Нейронное кодирование — это преобразование сигналов окружающей среды и внутренних сигналов тела в репрезентации этих сигналов в виде паттернов активности нейронов для создания модели реальности с целью адаптации и осуществления целенаправленных действий, сохранения целостности и нормального функционирования организма.
Так как формирование модели реальности и управление телом на её основе является главной задачей нервной системы , то расшифровку нейронного кода можно назвать центральной темой в нейронауке . Если мы достигнем понимания сути нейронного кода, то нам станет доступна внутренняя работа сознания в норме и при патологических отклонениях. Решение такой задачи является, возможно, наиболее важным направлением в науках о человеке, так как от работы мозга зависит течение всех жизненных процессов нашего организма.
Мозг — это высшее достижение эволюции природных информационных технологий в плане скорости и эффективности. Из этого следует, что из всех схем кодирования наиболее вероятным кандидатом на нейронный код является та, которая производит информацию (паттерны кода) наиболее эффективно.
Все современные модели рассматривают потенциал действия (спайк) нейрона как фундаментальный элемент языка мозга. Однако принципиальным является вопрос подхода к этому явлению. Хотя реальные потенциалы действия являются континуальными колебательными процессами и отличаются периодом, амплитудой и формой фазового портрета, ведущие модели рассматривают их как идентичные дискретные события.
Такие теории можно разделить на два класса: кодирование средней скоростью спайков и темпоральный код. Они формулируют вопрос следующим образом: либо информация содержится в количестве спайков за некоторое временное окно, либо в их точном местоположении на временной оси. В любом случае, все они представляют собой разные версии подсчета цепочек спайков ( англ. spike train).
Некоторые из этих моделей противоречат реальной эффективности и скорости работы мозга. Некоторые охватывают лишь часть наблюдаемых явлений и не могут объяснить другие. Данные модели существуют уже десятки лет, но не привели к расшифровке нейронного кода. Возможно, причина в неверности подхода к сути процесса нейронного кодирования. В последнее время появляются модели, отвечающие на вопросы, которые неразрешимы в рамках парадигм, относящихся к потенциалам действия нейронов как к одинаковым спайкам.
Стандартным подходом при изучении нейронного кода является поиск связи между входящим сигналом и реакцией нейронов, а также обратный процесс восстановления сигнала из наблюдаемой активности нейронов. Однако без модели самого кода такой анализ напоминает попытку прочтения или написания текста без знания грамматики. Возникает подобие замкнутого круга, когда для прочтения кода нужно его знать, а для того, чтобы его узнать, нужно читать.
Однако любой процесс расшифровки неизвестного кода строится на поисках определенных закономерностей и выявления их связи с кодируемым посланием. Иными словами, для прочтения нейронного кода нужно найти соответствие паттернов параметров сигналов паттернам активности нейронов.
Любой сигнал среды является колебательным энергетическим процессом с определенной амплитудой , частототой и развитием фаз во времени. Это две главные оси измерения сигнала: пространственная и временная. Соответственно и нейронный код должен обладать пространственными и временными характеристиками, создающими модель кодируемого сигнала.
По мере продвижения по иерархии технологической цепочки нервной системы от сенсоров на периферии к интегративным структурам коры головного мозга активность нейронов всё меньше связана с оригинальным сигналом. Это закономерный процесс, так как нейроны не отражают сигналы, а кодируют их, то есть создают репрезентации. Сознание — это не зеркало реальности, а моделирование реальности. Однако репрезентация все равно должна содержать в себе все те же оси измерения параметров. Таким образом нейронный код просто обязан быть сложной многомерной структурой. При этом информационная насыщенность должна сочетаться с эффективностью и скоростью.
Отражают ли эти требования предлагаемые модели кодирования? Этот вопрос должен быть «лакмусовой бумагой» для тестирования их адекватности реальным процессам в нервной системе.
Кодирование скоростью генерации потенциалов действия — это гипотеза, предполагающая, что информация о сигнале содержится в частоте срабатывания нейрона. Она появилась после проведения экспериментов Эдрианом и Зоттерманом в 1926 году. В этом простом опыте к мускулам подвешивали разные грузы. По мере увеличения веса количество импульсов иннервирующих мышцу нейронов также увеличивалось. Авторы пришли к выводу, что потенциалы действия являются дискретными событиями, и что их частота, а не индивидуальные параметры, являются основой нейронной коммуникации.
Идея была привлекательна своей простотой. При всей технической сложности регистрации спайков это была выполнимая задача даже на уровне возможностей оборудования прошлого века. Но кажущаяся простота обманчива, так как возникает много вопросов.
Даже на уровне периферии (сенсоры и эффекторы) частота возбуждения увеличивается, как правило, нелинейно с увеличением интенсивности стимула. Прямой связи между скоростью спайков и сигналом нет. К тому же последовательность потенциалов действия варьируется от испытания к испытанию. Поэтому для определения скорости применяются разнообразные произвольные алгоритмы усреднения. Сам термин «скорость стрельбы» (англ — firing rate) имеет несколько разных определений, которые относятся к различным процедурам усреднения, например, к среднему значению во времени или среднему значению за несколько повторений эксперимента. Даже если мы остаемся верны идее кода как средней скорости, применяемые методы усреднения могут совсем не соответствовать тому алгоритму, который используют сами нейроны.
Есть и еще одна проблема. Если мы хотим что-то усреднить, нам нужно знать временной интервал, в течение которого мы берем набор сэмплов для расчета. Если мы анализируем сигнал, повторяющийся со строгой периодичностью, то нам не составит труда определить время его периода и вычислить среднее значение. Но такой периодический сигнал не может создать информацию. Не удивительно, что нейроны не демонстрируют такую монотонность своих спайков.
Но если изменение не является строго периодическим, то выбор окна усреднения становится решающим. База для усреднения определяет конечный результат. Даже если мы примем гипотезу о том, что нейроны создают код средней скоростью стрельбы, мы должны понимать, что нейроны будут вычислять это среднее относительно длины цикла, который им известен. Но тогда нам нужно определить этот такт. Однако экспериментаторы измеряют количество спайков относительно произвольно выбранных временных окон. Типичные значения T = 100 мс или T = 500 мс, но продолжительность также может быть больше или меньше. Основное требование состоит в том, что во временном окне должно произойти хотя бы несколько спайков, чтобы получить средние значения.
Возникает замкнутый круг: мы не знаем является ли код действительно таким и не знаем, как это подтвердить или опровергнуть, потому что не знаем такт системы. В итоге, мы можем бесконечно произвольно усреднять, но это ничего не даст.
Но есть еще одна проблема, которая содержится в самой идее такого кода. Может ли код, состоящий из идентичных спайков, обеспечить наблюдаемую информационную насыщенность, скорость и эффективность работы нервной системы? Ответ на этот вопрос, к сожалению для приверженцев такой парадигмы, отрицателен. Такой код можно назвать темповым кодом. Однако вариация темпа не несет в себе достаточно информации для репрезентации сложного многопараметрового сигнала. К тому же, возникает дилемма: как закодировать быстрый сигнал, если нужно накапливать достаточное количество спайков, чтобы получить среднюю и извлечь какую-либо информацию из этой переменной.
Динамика огромного количества сигналов среды измеряется в миллисекундах, и в течение этих миллисекунд нейроны могут сработать только один или два раза. При таком количестве спайков невозможно кодировать сигнал их средней скоростью. Но есть и более быстрые сигналы. Например, летучая мышь способна к эхолокации с разрешением в микросекунды. Таким образом, временное окно измерения сигнала находится внутри одного спайка. Это полностью противоречит парадигме средней скорости.
Такой код неэффективен по всем параметрам. Он мало информативен и слишком медленный. Он требует создания множества спайков для кодирования простых параметров, то есть энергетически очень затратный. Именно поэтому он не соответствует реальности работы мозга. Однако эта модель до сих пор широко используется не только в экспериментах, но и в моделях нейронных сетей. В итоге, за прошедшие десятилетия накопилось огромное количество данных, но оно никак не приблизило нас к как расшифровке смысла кода.
Следующим кандидатом является кодирование, к котором информация содержится в точном местоположении спайков и межспайковых интервалов на временной оси. Так как эта модель снова рассматривает потенциалы действия как одинаковые дискретные события, то такую версию кода можно назвать ритмом чередования 1 и 0. Темпоральное кодирование позволяет последовательности 000111000111 означать что-то отличное от 001100110011, даже если средняя скорость срабатывания для обеих последовательностей одинакова. Такая модель была следующей очевидной идеей и альтернативой неэффективному и медленному варианту кода средней скорости.
Предположение о том, что нейронный код двоичный, а не состоит только из спайков, значительно повышает емкость кода и придает правдоподобия модели. Но встает всё тот же вопрос соотнесения информационной ёмкости кода и реальной скорости работы мозга, который успевает закодировать сложный многопараметровый сигнал в течение одного-двух спайков. У мозга нет времени на построение длинной бинарной цепочки, которая могла бы содержать в себе всю информацию. В этом он принципиально отличается от искусственных цифровых систем. При всей огромной скорости их процессоров, которая на порядки выше частот мозга , они не могут сравниться с ним в производительности, скорости и энергетической эффективности. Проблема в том, что им нужно обрабатывать длинные двоичные коды .
К тому же снова встает вопрос о такте системы. Два нуля кода — это пауза вдвое длиннее одного нуля. Но как определить, что межспайковая пауза означает два нуля или один, если мы не знаем шкалу времени изучаемой системы? Измерение паузы по внешним часам дает нам много данных, но ничего не говорит о том, сколько нулей находится в этой конкретной паузе и как их соотносить с единицами спайков. Иными словами, мы не можем определить означает ли активность нейрона 0001 или 001.
Для реального качественного анализа необходимо нормализовать данные системы по её собственному времени, по её частотам. Тогда мы сможем выразить наш анализ в любой единице измерения. Поиск этой фундаментальной частоты, в качестве основы для нормализации, вероятно, имеет первостепенное значение при попытке разгадать код мозга вне зависимости от того, какую модель кода мы тестируем, так как временной параметр остается в любом случае.
Некоторые исследователи считают, что есть и темповый, и темпоральный код, и есть нейроны-переводчики между этими кодами. Эта идея, конечно, примиряет две концепции, но не решает проблемы, присущие обеим.
Эта версия кода направлена на выход из тупиков предыдущих моделей. Она показывает, что для подсчета спайков нужна привязка к определенной временной метке и предлагает поиск такой системы отсчета в частотах мозга.
Эксперименты показывают, что нейроны кодируют сигналы среды, соотнося время активации с фазой текущих колебаний нейронной сети. Таким образом образуется определенная последовательность спайков, соотносимая с общей временной шкалой сети. Принципиальным моментом является определение несущей частоты, которую можно было бы с уверенностью назвать системой отсчета.
Исследователи рассматривают такую модель как дополнение к коду скорости стрельбы. Они продолжают считать спайки одинаковыми импульсами и видят информацию только в ритмической структуре активации нейронов.
Есть версии кода, которые говорят о том, что активность отдельных нейронов не содержит никакой информации и смысл нужно искать в паттернах объединенной активности. В таких моделях считается, что нейроны стреляют в случайном порядке с распределением Пуассона и такой хаос создает порядок в виде популяционного кода. При этом модель ничего не говорит о том, каков механизм работы и правила такого кода.
Тонкие методы измерения с помощью вживляемых электродов и подробное изучение темпоральной структуры самих спайков и межспайковых интервалов показывают, что она не носит характер распределения Пуассона и каждый из атрибутов стимула изменяет не только абсолютное количество спайков, но и их темпоральный паттерн. Несмотря на огромную вариативность активности нейронов, последовательности спайков очень точны. Эта точность необходима для передачи информации с помощью кода высокого разрешения. Кажущаяся хаотичность активации происходит из непонимания сути кода.
Такую модель можно назвать реакцией на то, что десятилетия попыток рассшифровать нейронный код путем подсчета спайков и поиска смысла в скорости или темпоральной структуре их последовательностей не привели к результату.
В целом популяционная версия кода справедливо указывает на то, что репрезентации сигналов являются результатом активности многих нейронов. Каждый нейрон занимает свое место в процессе формирования смыслов и обладает специализацией как фильтр по преобразованию определенных параметров сигнала. Однако встает вопрос о том, как паттерны активности каждого нейрона соединяются в общую репрезентацию сигнала со всеми параметрами и как репрезентации отдельных сигналов сливаются в единую и когерентную модель реальности, сохраняя при этом свою индивидуальность. В нейронауке это называется «проблемой соединения» (англ — ).
Некоторые модели популяционного кода описывают этот процесс математически как сумму векторов всех нейронов, участвующих в кодировании данного сигнала. Векторное кодирование — это пример простого усреднения. Есть более сложные математические модели, использующие понятия теории вероятности. Однако общей проблемой таких моделей является отсутствие объяснения физического механизма, который мог бы осуществлять наблюдаемое единство создаваемой мозгом модели реальности с сохранением индивидуальности репрезентаций сигналов.
Такая модель кодирования утверждает, что корреляции между потенциалами действия в цепочке спайков могут нести дополнительную информацию помимо простого определения времени спайков. Например, эксперименты показывают, что звуковой сигнал определенной частоты вызывает увеличение количества коррелированных спайков в слуховой коре и не влияет на среднюю частоту спайков. Очевидно, что соотношение потенциалов действия между собой несет информацию в общем процессе кодирования. Эту идею можно назвать движением от кода средней скорости в сторону более адекватной модели, говорящей об информационной насыщенности пространственно-темпоральных паттернов активности нейронов. Однако её нельзя назвать самостоятельной и сформированной моделью нейронного кода.
Понятие разреженности нейронного кода может относиться к разреженности во времени или к разреженности активной популяции нейронов. Однако это нельзя назвать моделью кода, так как является просто констатацией того, что «нейроны имеют свойство быть редкими, и в смысле небольшого числа активных нейронов в определенный момент времени, и в смысле того, что индивидуальный нейрон может стрелять достаточно редко в течение определенного времени».
Экспериментально разреженность кодирования наблюдались во многих системах, включая зрение , слух , осязание и обоняние. Данный факт еще раз подчеркивает неадекватность модели средней скорости, так как она подразумевает необходимость большого количества спайков для получения даже небольшого информационного набора.
Каковы задачи нейронов при обработке сигналов внешней и внутренней среды? Во-первых, нейроны должны создавать информационно эффективный код. Во-вторых, нейроны должны создавать энергетически эффективный код. Эти требования ведут к разреженности кода в смысле малого количества элементов в быстром временном окне и малого набора базовых элементов кода, которые могут кодировать сложную информацию своими сочетаниями. Из этого следует вывод, что каждый элемент должен быть информационно насыщенным. Иными словами, нейронный код должен сочетать в себе разреженность и насыщенность. Это не взаимоисключающие, а взаимодополняющие требования.
Возникает вопрос: может ли спайк нейрона быть информационно насыщенным, если это дискретное событие, не имеющее внутренних характеристик? В такой постановке вопрос становится риторическим и ответ на него очевидно отрицательный. К сожалению, все вышеуказанные модели исходят из допущения о том, что потенциалы действия нейронов одинаковы. Но возникает следующий вопрос: так ли это на самом деле? Более того, возникает вопрос: являются ли потенциалы действия спайками (англ. spike – острый шип)?
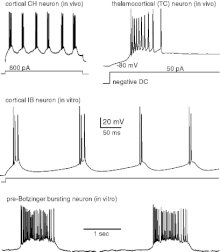
Стандартное изображение активности нейронов выглядит как шипы, разложенные с разной плотностью по оси времени:

Может быть, потенциалы действия действительно такие ровные палочки? На самом деле нет. Так их упрощают в исследованиях, чтобы сделать удобными для модели. Но это не первый и, наверное, не последний раз в науке, когда реальность подстраивают под теорию, а не меняют модель, когда она противоречит реальности.
«Спайк добавлен вручную в эстетических целях и для одурачивания читателя, чтобы он поверил, что нейрон производит спайк ... Все спайки принимаются как одинаковые по размеру и продолжительности ... Несмотря на все недостатки, модель интегрируй-стреляй является приемлемой жертвой для математика, который хочет доказать теоремы и вывести аналитические формулы. Однако такая модель может быть просто потерей времени».
Действительно, за десятилетия в рамках такого подхода написаны сотни тысяч научных публикаций и созданы сотни теорий и моделей нейронного кода, основанных на идеализации и рисовании одинаковых спайков. Возможно, это действительно потеря времени, потому что такие модели не соответствуют реальности и приводят к неразрешимым тупикам.
Чтобы понять, отражает ли эта картина реальность или нет, мы должны обратиться к временному уровню самого нейрона. Если увеличить разрешение по оси времени, то станет видно, что нейроны не стреляют острыми шипами, а вибрируют мягкими волнами.
После десятилетий рассматривания дискретных единиц там, где на самом деле волны, мы снова возвращаемся к вопросу: «Какова структура нейронного кода, позволяющая такие высокие уровни передачи информации? ... Нам нужна теория таких вычислений ... Природа построила вычислительную машину удивительной точности и адаптивности ... История началась, более или менее, с открытия Адриана, что спайки — это единицы, из которых должно строиться наше восприятие. Мы приходим к мысли, что каждая из этих единиц вносит определенный и измеряемый вклад в это восприятие. Каждый спайк, который так часто усредняется вместе с соседями, заслуживает большего уважения».
Гипотеза симфонического нейронного кода предполагает, что нейронный код не цифровой, а аналого-цифровой. Это означает, что активность нейрона содержит в себе параметры каждого потенциала действия как аналоговую (волновую) составляющую спайка, который одновременно является и дискретной единицей кода.
Данная теория использует аналогию с музыкальным кодом (нотной записью). В этом смысле каждый потенциал действия нейрона является нотой музыки мозга, т.е. обладает индивидуальными характеристиками формы волны (периодом, амплитудой, фазовым портретом). Индивидуальные ноты составляют паттерн активности данного нейрона с четкой пространственно-временной организацией, позволяющей встраиваться в общую музыку мозга с её мелодиями (частотный паттерн), ритмами (фазовый паттерн) и гармониями (одновременное существование разных паттернов). Информационная насыщенность каждой ноты (потенциала действия) и каждой паузы (потенциала покоя) очень высокая. Таким образом сложная информация может содержаться в цепочке из очень небольшого количества последовательностей спайк/пауза и даже в пределах одного спайка. Благодаря этому вся система в целом обладает огромной вычислительной мощностью, эффективностью и скоростью.
В некотором смысле эта модель объединяет многие предыдущие гипотезы. Она показывает, что скорость генерации спайков имеет место в общей структуре кода. Но, как и в музыке, темп не имеет самостоятельного значения. Модель подчеркивает важность темпоральной (ритмической) структуры, несущей большую информационную нагрузку. Это, безусловно, модель кодирования фазой, поскольку динамика активации/деактивации и внутренняя структура фазового портрета каждого потенциала действия позволяют кодировать сигналы даже во временном окне фазового сдвига в пределах одного цикла активации. Это модель популяционного кода, которая рассматривает нейроны не как бессмысленные или шумные компоненты системы, которые создают порядок из хаоса каким-то волшебным способом, а как участников оркестра мозга, состоящего из миллиардов музыкантов, играющих единую симфонию, в которой каждая партия имеет собственное значение. Это, безусловно, корреляционное кодирование, поскольку каждая партия расположена в контексте всей структуры. Это и модель разреженного кода, поскольку симфония может состоять из потенциально бесконечных комбинаций малого набора базовых нот (элементов кода), а небольшое количество музыкантов (нейронов) может принимать участие в создании сложной комбинации. В то же время это модель насыщенного кодирования, поскольку каждая нота несет высокую информационную нагрузку.
Использование музыкальной терминологии не метафора, а физическая аналогия. Физика процесса нейронного кодирования основана на колебательных и волновых явлениях, так же как и создание звуков, которые мы называем музыкой. Кардинальный сдвиг парадигмы состоит в том, что нейрон рассматривается не как производитель одинаковых выстрелов, а как осциллятор со сложным фазовым портретом. Такая модель отдает дань уважения каждому потенциалу действия. Вся тонкая логистика организации и кинетика процессов на внутриклеточном и межклеточном уровнях предназначена для создания параметров колебаний нейронов. Модель дает подробное физическое, математическое и технологическое описание процесса нейронного кодирования, которое объясняет информационную, темпоральную и энергетическую эффективность мозга.
Более того, подход к процессу кодирования как к взаимодействию осцилляторов с разными параметрами позволяет совершенно иначе взглянуть на проблему соединения и показать реальный физический механизм частотного и фазового сцепления, создающий единую симфонию сознания как гармоническую структуру с сохранением индивидуальных характеристик каждой репрезентации. Такая модель также позволяет нам рассматривать патологии, которые в настоящее время считаются загадочными «психическими расстройствами» (например, аутизм и шизофрения ), как конкретные нарушения в кодировании сигналов мира и создании адаптивной модели реальности.
Мы должны оказывать должное уважение индивидуальным потенциалам действия и всей тонкой динамике пространственно-временных паттернов нейронной активности, которые составляют суть процесса кодирования. С одной стороны, это выводит нас из бесконечных блужданий по лабиринту и дает нам « нить Ариадны », чтобы выбраться на свет. С другой стороны, это имеет последствия, которые нельзя назвать тривиальными по двум причинам. Во-первых, это противоречит господствующим теориям и требует смены парадигмы, что непросто. Во-вторых, резко увеличивается пространство параметров описания. «Ясно, что невозможно прыгнуть от подсчета спайков к 'полному' анализу». Исследователи не готовы «прыгать». Гораздо удобнее оставаться в обычном режиме подсчета спайков, который используется уже почти сто лет.
Сказать, что это была пустая трата времени, неправильно, так как в научных исследованиях любой результат, даже отрицательный, имеет значение. Расшифровка любого кода — долгий путь, на котором есть тупиковые направления. Но единственный выход — осознать, что это тупик, и изменить маршрут. Так или иначе, многие технологии для изучения мозга были разработаны на этом пути. Традиционные инструменты (например, ЭЭГ , МЭГ , ФМРТ ) не подходят для расшифровки кода, поскольку в настоящее время они не обладают необходимым пространственно-временным разрешением и не измеряют активность нейронов напрямую. Другие технологии (например, , метод локальной фиксации потенциала ) лучше подходят для этой задачи, но имеют свои недостатки. Некоторые новые технологии, такие как оптогенетика , позволяют измерять и даже контролировать активность отдельных нейронов.
Давно назревшая смена парадигмы ведет к сложным концептуальным и технологическим последствиям для процесса экспериментальных исследований. Нейронный код оказался не таким простым, как мы думали, потому что мир, который он кодирует, непростой. Мозг сложен, но это означает, что он может анализировать сам себя. Наш разум способен расшифровать собственный код. Для этого он должен создать правильную модель, показывающую выход из «лабиринта». Если произойдет концептуальный прорыв, технология последует за ним.
Расшифровка нейронного кода— широкомасштабная задача по своей сложности и значимости. Трудно переоценить перспективы, которые откроет для нас возможность читать и писать нейронный код. Они касаются лечения не только психических и неврологических расстройств, но и всех заболеваний организма, от трех основных (сердечно-сосудистые, рак и диабет) до менее серьезных, но все же влияющих на качество жизни. Это также откроет новую перспективу для развития технологий искусственного интеллекта и их интеграции с естественными технологиями мозга.