Interested Article - Миксомицеты

- 2021-06-05
- 1
Слизевики , или миксомицеты ( лат. Myxomycetes ), — класс настоящих слизевиков ( Mycetozoa ), включающий более 800 видов организмов, крупнейшая группа среди амёбозоев . Встречаются повсеместно от тропиков до полярных регионов .
История изучения
Первые зафиксированные наблюдения миксомицетов относятся к XVII веку. Томас Панковиус упоминает ликогалу древесинную в своём труде «Herbarium Portatile, oder behendes Kräuter- und Gewächsbuch» 1654 года как «быстрорастущий гриб» ( лат. Fungus cito crescentes ) . Из-за сходства с гастеромицетами миксомицеты в Systema mycologicum были отнесены Элиасом Магнусом Фрисом к подклассу Myxogastres класса Gasteromycetes. В 1833 году Генрих Фридрих Линк выделил миксомицетов в отдельный класс грибов, дав ему название Myxomycetes .
В 1859 году в своей работе «Die Mycetozoen» Генрих Антон де Бари показал отличия миксомицетов от грибов и ввёл термин Mycetozoa («грибо-животные»). В настоящее время установлено, что миксомицеты относятся к группе Amoebozoa, в равной степени близкой грибам и животным. Тем не менее, их изучением по традиции занимаются преимущественно микологи .
Жизненный цикл и строение


Жизненный цикл миксомицетов начинается с гаплоидных спор . Из них, в зависимости от влажности окружающей среды, формируются или жгутиковые зооспоры , или амёбоидные . Промежуточную стадию между ними принято называть мастигамёбами. При неблагоприятных условиях зооспоры и миксамёбы способны переходить в состояние покоя, образуя микроцисту.
В результате слияния двух гаплоидных клеток образуются диплоидные клетки. Из них путём множественных синхронизированных митозов развивается плазмодий — многоядерная, сложно дифференцированная клетка, покрытая только плазматической мембраной . Плазмодий обладает отрицательным фототаксисом и положительными гидро- , рео- и , то есть стремится к затенённым влажным местам с большим количеством питательных веществ. При неблагоприятных условиях плазмодии могут формировать покоящиеся стадии — и/или склероции .
При определённых условиях плазмодии переходят к формированию спороношений. Все таксисы меняются на обратные. В результате получаются спорофоры , которые содержат гаплоидные споры, образовавшиеся посредством мейоза . На этом жизненный цикл замыкается .
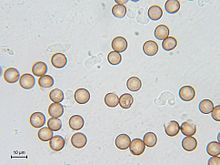
Споры
После разрушения спорофора споры претерпевают период покоя, который иногда может длиться несколько десятков лет . Споры у большинства миксомицетов сферические, реже эллипсоидальные и яйцевидные. Цвет споровой массы и отдельных спор в проходящем свете — важные диагностические признаки. Оболочка спор может быть снабжена такими структурами как бородавочки (с закруглённой вершиной) и шипы (с заострённой вершиной), но бывают и гладкие споры .
Миксамёбы и зооспоры
Миксамёбы лишены жёсткой клеточной стенки и способны перемещаться по субстрату с помощью псевдоподий . При повышенной влажности они способны за 1,5—2 часа переходить в стадию зооспоры и обратно. В условиях уменьшения влажности и недостатка питания миксамёбы и зооспоры могут образовывать микроцисты с плотными полисахаридными оболочками. При наступлении благоприятных условий микроцисты могут прорасти даже после длительного замораживания и высушивания. При этом оболочка растрескивается и из неё выходят новые миксамёбы или зооспоры .
Плазмодии

Способность к образованию плазмодиев является уникальной чертой миксомицетов. Они представляют собой многоядерную трофическую стадию, способную к амёбоидному движению. Скорость плазмодия может достигать 0,1—0,4 мм в минуту. В плазмодии наблюдаются ритмичные токи цитоплазмы .
Различают три основных морфологических типа плазмодиев :
- Протоплазмодий — имеет микроскопические размеры. В нём отсутствуют ритмические токи цитоплазмы. Встречается у представителе порядков Echinosteliales и Cribrariales.
- Афаноплазмодий — характеризуется отсутствием развитого слизевого чехла и наличием особой стадии «коралла» перед началом формирования спорофоров. Характерен для представителей порядка Stemonitidales. Наиболее устойчив к обезвоживанию .
- Фанероплазмодий — обладает слизистым чехлом и ритмичными токами цитоплазмы. Часто бывает ярко окрашен. Наиболее характерен у представителей порядка Physarales, но имеется и у некоторых представителей Cribrariales.
Иногда выделяют ещё один тип плазмодия, занимающий по морфологии промежуточное положение между афаноплазмодием и фанероплазмодием. Он характеризуется наличием токов цитоплазмы, но имеет меньшие размеры и более слабую пигментацию. Такой тип плазмодия характерен для представителей порядка Trichiales .
В неблагоприятных условиях плазмодии могут образовывать склероции или фрагментироваться на отдельные сгустки цитоплазмы, покрытые плотной оболочкой — сферулы .
Спороношения
Выделяют четыре основных типа спороношений миксомицетов, между которыми бывают промежуточные варианты :
- Плазмодиокарпы — наиболее простой тип спорофоров. Образуется при созревании плазмодия, покрытого общей оболочкой, почти без изменения его конфигурации. По форме представляется собой короткие или длинные шнуры, вытянутые ветвящиеся или сетчатые образования.
- Спорангии — шаровидные, овальные, колончатые или другие спороносные структуры, формирующиеся в результате дробления плазмодия. Могут быть сидячими или на ножках.
- Псевдоэталии образуются при срастании спорангиев боковыми стенками и потере ими индивидуальности, с сохранением возможности подсчитать количество спорангиев.
- Эталии — тип спорофоров, образующийся при срастании спорангиев с полной потерей их индивидуальности. Представляют собой подушковидные образования, покрытые общей оболочкой (кортексом).
- Типы спорофоров
-
 Плазмодиокарп
Плазмодиокарп -
 Спорангии
Спорангии -
 Эталий Reticularia lycoperdon
Эталий Reticularia lycoperdon -
 Псевдоэталий Tubifera ferruginosa
Псевдоэталий Tubifera ferruginosa
Экология
Трофические стадии миксомицетов, такие как миксамёбы, представляющие собой доминирующую группу почвенных простейших , питаются бактериями . Бо́льшую часть информации об экологии этих организмов получают при изучении их спорофоров, из-за чего исследователи в основном имеют дело с данными об их расселительных стадиях .
На основании изучения особенностей распространения спорофоров выделяют следующие эколого-трофические группы :
- Лигнофильные — встречаются на гниющей древесине. Самая большая группа, включающая до 70 % известных миксомицетов. Большинство из них обладают макроскопическими размерами. Пики спороношений наблюдаются с середины лета до поздней осени.
- Кортикальные — обитают на коре живых деревьев и кустарников. Обычно небольшие (от 100 мкм до 1—2 мм). Доминируют в пустынях и местах с высокой антропогенной нагрузкой.
- Подстилочные и почвенные — питаются в верхнем почвенном слое. Спороношения образуют на растительных остатках. Видовой состав в разных слоях листового опада неоднороден .
- Копрофильные — обитают на выветренном помёте растительноядных млекопитающих и птиц . Наибольшего разнообразия достигают в зонах аридного климата . За всё время исследования было обнаружено 114 видов копрофильных миксомицетов, 16 из которых были специфичны для данного типа субстрата . Представителей этой группы редко находят в полевых условиях. Основным методом их изучения является метод «влажной камеры». В отличие от грибов, большинство копрофильных миксомицетов не проходят через желудочно-кишечный тракт животных, а заселяют субстрат спустя продолжительное время. В то же время, споры некоторых видов обладают толстыми стенками, что может быть адаптацией к прохождению через пищеварительную систему .
- Бриофильные — обитают на мхах . Часто ассоциированы с микроскопическими водорослями . Являются самой малочисленной группой, включающей менее 5 % известных видов. Большинство представителей имеют микроскопические размеры. Появляются поздней осенью.
- Нивальные — развиваются весной на растительных остатках возле тающего снега. Наибольшего разнообразия достигают в альпийских и субальпийских районах северного полушария , но могут встречаться и в горах южного полушария и на равнинах . Для развития спороношений нивальных миксомицетов необходимо наличие снежного покрова в осенние месяце для предотвращения промерзания почвы .
Один и тот же вид миксомицетов может обитать на разных типах субстрата и потому может быть отнесён к разным эколого-трофическим группам.
Таксономия
Классическая система
Класс Myxomycetes включает пять порядков.
Echinosteliales
Порядок включает 2 семейства — и . Эхиностелиевые рассматриваются как наиболее близкая к группа миксомицетов. Это заключение носит предварительный характер, так как ультраструктура жгутиков зооспор некоторых протостелиевых и видов рода отличается. В то же время наблюдается значительное сходство в строении одноядерных миксамёб протостелиевых и протоплазмодия эхиностелиевых, а также сходство морфологии плодовых тел у представителей обеих групп. Очевидно, для выяснения положения рода требуются дополнительные молекулярно-биологические исследования.
Liceales

Порядок включает 3 семейства: , , , для представителей которых характерно отсутствие капиллиция . Однако этот признак встречается также у рода из порядка . Некоторые виды рода обладают сходными признаками с ( Trichiales ) и ( Liceales ), что ставит под сомнение гомогенность рода в современном понимании его объёма.
Trichiales
Порядок включает 2 семейства: и Trichiaceae . Границы между родами не всегда чётко определены, так же как и их положение в системе. У видов первого нити капиллиция сплошные, у второго — полые. В пределах этого семейства усложнение капиллиция происходило как за счёт усложнения системы капиллиция в целом, так и её отдельных элементов. Наиболее просто устроен капиллиций у представителей рода Perichaena , состоящий из редко ветвящихся трубочек. Очевидно, такой тип можно рассматривать как плезиоморфный признак. Отсутствие чётко выраженного протока в нитях капиллиция у видов рода , возможно, потребует пересмотра положения этого рода в семействе Trichiaceae . Следующим этапом усложнения капиллиция считается появление почти неорнаментированных и не связанных с перидием элатер, как у представителей рода . У видов рода заметно усложнение орнаментации нитей капиллиция за счёт образования на них шипиков и спиральных утолщений. Связь капиллиция с перидием и образование сети нитей, а также их способность вытягиваться при созревании спорангия и изменении влажности, что приводит к растрескиванию перидия, является следующим этапом дифференциации этой структуры. Подобный тип характерен для родов и . Основное отличие между ними заключается в различной орнаментации отдельных трубочек капиллиция.
Stemonitales
Порядок Stemonitales включает единственное семейство Stemonitidaceae . В пределах семейства имеются роды с весьма размытым ареалом. Это — , Stemonitis , , , .
Physarales
Порядок включает два семейства и . На основании наблюдений за развитием и в результате электронно-микроскопических исследований роды , , были перемещены из Stemonitales в Physarales . Что касается родов Diachea и Leptoderma , то развитие спорофора по субгипоталлическому типу и наличие у видов этих родов указывают на их принадлежность к порядку Physarales .
Филогенетическая система
В 2019 году была предложена система миксомицетов, основанная на последовательностях гена малой субъединицы рРНК , и отражающая их филогенетические связи :
-
Подкласс
Lucisporomycetidae
Leontyev, Schnittler, S.L. Stephenson, Novozhilov & Shchepin — включает представителей со светлыми спорами без
меланина
.
-
Надпорядок
Cribrariidia
-
Порядок
Cribrariales
T. Macbr.
- Семейство Cribrariaceae Corda
-
Порядок
Cribrariales
T. Macbr.
-
Надпорядок
Trichiidia
-
Порядок
Liceales
E. Jahn
- Семейство Liceaceae Chevall.
-
Порядок
Reticulariales
Leontyev, Schnittler, S.L. Stephenson, Novozhilov & Shchepin
- Семейство Reticulariaceae Chevall.
-
Порядок
Trichiales
Chevall.
- Семейство Trichiaceae Chevall.
- Семейство Dianemataceae T. Macbr.
-
Порядок
Liceales
E. Jahn
-
Lucisporomycetidae
incertae sedis (светлоспоровые роды с неясным систематическим положением)
- Hochg. & Gottsb.
- Morgan
- G. Lister
- Novozh., Hoof & Jagers
-
Надпорядок
Cribrariidia
-
Подкласс
Columellomycetidae
Leontyev, Schnittler, S.L. Stephenson, Novozhilov & Shchepin — включает преимущественно представителей с тёмной споровой массой, окраска которой определяется содержанием меланина.
-
Надпорядок
Echinosteliidia
-
Порядок
Echinosteliales
G. W. Martin
- Семейство Echinosteliacea Rostaf. ex Cooke
-
Порядок
Echinosteliales
G. W. Martin
-
Надпорядок
Stemonitidia
-
Порядок
Clastodermales
Leontyev, Schnittler, S.L. Stephenson, Novozhilov & Shchepin
- Семейство Clastodermataceae Alexop. & T. E. Brooks
-
Порядок
Meridermatales
Leontyev, Schnittler, S.L. Stephenson, Novozhilov & Shchepin
- Семейство Leontyev, Schnittler, S.L. Stephenson, Novozhilov & Shchepin
-
Порядок
Physarales
T. Macbr.
- Семейство Didymiaceae Rostaf. ex Cooke
- Семейство T. Macbr.
- Семейство Physaraceae Chevall.
-
Порядок
Stemonitidales
T. Macbr.
- Семейство Rostaf. ex Cooke
- Семейство Stemonitidaceae Fr.
-
Порядок
Clastodermales
Leontyev, Schnittler, S.L. Stephenson, Novozhilov & Shchepin
-
Columellomycetidae
incertae sedis (темноспоровые роды с неясным систематическим положением)
- Fr.
- Reinhardt & L. S. Olive
- G. Lister
- Hertel
- M. Blackw. & Alexop.
- H. W. Keller
- Kuntze
-
Надпорядок
Echinosteliidia
Примечания
- (англ.) на сайте MycoBank . (Дата обращения: 18 февраля 2020) .
- ↑ , с. 16—20.
- Steven L. Stephenson, Yuri K. Novozhilov, Martin Schnittler. (англ.) // Journal of Biogeography. — 2000-05. — Vol. 27 , iss. 3 . — P. 741–754 . — ISSN . — doi : .
- Steven L. Stephenson, Rodney D. Seppelt, Gary A. Laursen. (англ.) // Antarctic Science. — 1992-12. — Vol. 4 , iss. 4 . — P. 431–432 . — ISSN . — doi : .
- ↑ Dmitry V. Leontyev, Martin Schnittler, Steven L. Stephenson, Yuri K. Novozhilov, Oleg N. Shchepin. // Phytotaxa. — 2019-03-27. — Т. 399 , вып. 3 . — С. 209 . — ISSN . — doi : . 13 января 2022 года.
- , с. 11.
- , с. 5.
- ↑ , с. 7—15.
- Ernest C. Smith. // Mycologia. — 1929-11. — Т. 21 , вып. 6 . — С. 321 . — doi : . 1 января 2022 года.
- Eugene W. Elliott. (англ.) // Mycologia. — 1949-03. — Vol. 41 , iss. 2 . — P. 141–170 . — ISSN . — doi : .
- , с. 23—25.
- Collins O. R. (англ.) // Botanical Review. — 1979. — Vol. 45 , no. 2 . — P. 145-201 . 1 января 2022 года.
- Steven L. Stephenson, Anna Maria Fiore-Donno, Martin Schnittler. (англ.) // Soil Biology and Biochemistry. — 2011-11. — Vol. 43 , iss. 11 . — P. 2237–2242 . — doi : . 30 июля 2020 года.
- M.F. Madelin. (англ.) // Transactions of the British Mycological Society. — 1984-08. — Vol. 83 , iss. 1 . — P. IN1–19 . — doi : . 25 июня 2018 года.
- Aw Rollins. // Mycosphere. — 2012-09-08. — Т. 3 , вып. 5 . — С. 543–549 . — doi : . 1 января 2022 года.
- ↑ Uno Eliasson. (англ.) // Fungal Diversity. — 2013-03. — Vol. 59 , iss. 1 . — P. 85–90 . — ISSN . — doi : .
- Y. K. Novozhilov, M. Schnittler, D. A. Erastova, M. V. Okun, O. N. Schepin. (англ.) // Fungal Diversity. — 2013-03. — Vol. 59 , iss. 1 . — P. 109–130 . — ISSN . — doi : .
- Steven L. Stephenson, John D. L. Shadwick. (англ.) // Australian Journal of Botany. — 2009. — Vol. 57 , iss. 2 . — P. 116 . — ISSN . — doi : .
- Ерастова Д. А. , Новожилов Ю. К. // Микология и фитопатология. — 15. — Т. 49 , № 1 . — С. 9—18 . — ISSN .
- , с. 347—353.
Литература
- Гмошинский В. И. , Дунаев Е. А. , Киреева Н. И. Определитель миксомицетов Московского региона. — М. : АРХЭ, 2021. — 388 с. — ISBN 978-5-94193-089-0 .
- Новожилов Ю. К. Определитель грибов России: Отдел Слизевики. — СПб. : Наука, 1993. — 288 с. — ISBN 5-02-026625-0 .
- Fiore-Donno A.M., Berney C., Pawlowski J., Baldauf S.L. higher-order phylogeny of plasmodial slime molds (Myxogastria) based on elongation factor 1-A and small subunit rRNA gene sequences (англ.) // J. Eukaryot. Microbiol. : journal. — 2005. — Vol. 52 , no. 3 . — P. 201—210 . — doi : . — .
- Kirk P. M., Cannon P. F., Minter D. W., Stalpers JA. (неопр.) . — Wallingford: CABI, 2008. — С. . — ISBN 0-85199-826-7 .
- Baldauf S.L., Doolittle W.F. (англ.) // Proc. Natl. Acad. Sci. U.S.A. : journal. — 1997. — October ( vol. 94 , no. 22 ). — P. 12007—12012 . — doi : . — . — PMC .
- Stephenson S. L. , Rojas C. Myxomycetes: biology, systematics, biogeography and ecology (англ.) . — 2nd ed. — Elsevier: Academic Press, 2021. — 600 p. — ISBN 9780128242827 .
|
|
|
|---|---|
| Таксономия |
|
- 2021-06-05
- 1



