Список эпизодов телесериала «Вавилон-5»
- 1 year ago
- 0
- 0

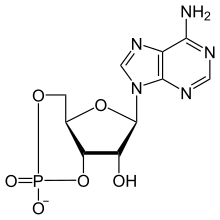
PTEN (сокр. от англ. p hosphatase and ten sin homolog deleted on chromosome 10 ) — фосфатаза с двойной субстратной специфичностью, продукт гена PTEN . Субстратами этой фосфатазы могут быть как белки , так и фосфатидилинозитол-3-фосфаты . PTEN катализирует отщепление фосфатной группы в положении 3D инозитольного кольца фосфатидилинозитол-3-фосфатов, лишая их таким образом функций вторичных посредников при передаче сигнала в клетке . Эта фосфатаза является одним из немногих негативных регуляторов PI3K/AKT/mTOR-сигнального пути , что делает её антионкобелком . Ген PTEN часто бывает мутирован при различных типах злокачественных опухолей .
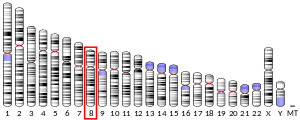
В 1980-х годах было обнаружено, что в ряде злокачественных опухолей головного мозга , мочевого пузыря и предстательной железы происходит частичная или полная потеря 10-й хромосомы . Однако только в 1997 году новый ген-супрессор опухолей PTEN был картирован в локусе 10q23.3 и было установлено, что он потенциально кодирует тирозиновую фосфатазу . Было установлено, что этот ген часто претерпевает мутации при различных видах спорадического рака, а также в случае наследственной предрасположенности к нему .
Последующие эксперименты на мышах, у которых этот ген был нокаутирован , показали важную роль PTEN в подавлении роста опухолей некоторых тканей, в работе сердца и мозга , метаболизме глюкозы , а также в дифференцировке и пролиферации Т- и В-лимфоцитов .
Ген PTEN имеет размер 105 тысяч пар оснований и содержит 9 экзонов .
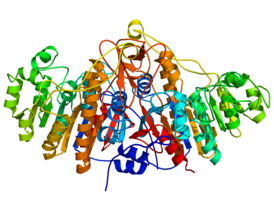
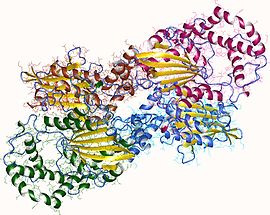
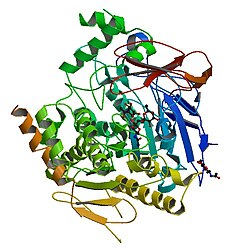
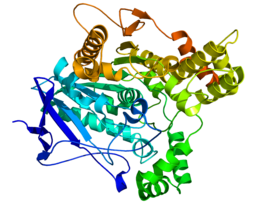
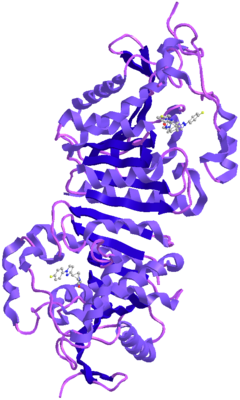
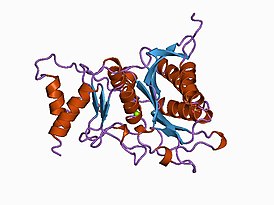
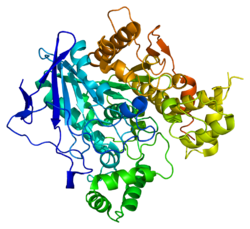
Фосфатаза PTEN состоит из 403 аминокислотных остатков и содержит аминокислотный мотив ( I / V ) H C X A G XX R ( S / T )G (где Х — любая аминокислота), характерный для тирозиновых протеинфосфатаз и фосфатаз двойной субстратной специфичности. PTEN содержит следующие структурные компоненты: короткий N-концевой домен связывания фосфатидилинозитол-4,5-бисфосфата (PBD, 1—13 а. о.), фосфатазный домен (14—185 а. о.), домен C2 (190—350 а. о.), C-концевой домен (351—400 а. о.) и мотив, связывающий PDZ-домен (401—403 а. о.) .
N-концевой фрагемент PTEN имеет голомогическое сходство по аминокислотной последовательности с актинсвязывающим белком тензином 1 (TNS1), откуда эта фосфатаза и получила своё название. Фосфатазный домен состоит из пятицепочечного β-листа , окружённого двумя α-спиралями с одной стороны и четырьмя — с другой. Домен C2 представляет собой β-сэндвич из двух антипараллельных β-листов с двумя короткими α-спиралями между этими листами. Этот домен отвечает за связывание липидов и удержание молекулы в мембране. С-концевой домен содержит PEST-последовательность и отвечает за регуляцию стабильности белка. Мутации, являющиеся причиной злокачественного перерождения клеток , обнаруживаются во всех доменах PTEN, что говорит об их важности для антионкогенной функции белка .
Уникальная фосфатидид-3-фосфатазная активность PTEN делает её одним из ключевых супрессоров опухолей в организме. PTEN катализирует отщепление фосфатной группы в положении 3D инозитольного кольца фосфатидилинозитол-3-фосфатов, тормозя передачу сигнала по PI3K/AKT/mTOR-сигнальному пути. К белковым субстратам PTEN относятся FAK, ETS2, Sp1, PDGFR . Нормальная работа PTEN необходима для контроля за пролиферацией клеток и их внедрением в соседние ткани. Делеция этого гена приводит к избытку андрогенов и дисфункции яичников у мышей .
В соответствии с антионкогенной функцией PTEN, нарушение её нормальной работы приводит к формированию опухолей . Мутации в гене PTEN ассоциированы с несколькими наследственными синдромами , характеризующимися развитием множественных доброкачественных опухолей в различных органах и тканях. К таким синдромам относятся синдром Коудена и болезнь Лермитт-Дуклос. Кроме того, мутации или биохимическая инактивация PTEN обуславливают предрасположенность к ряду онкологических заболеваний, таких как рак простаты , эндометрия и глиома . В экспериментах на мышах обнаружено что повысив с помощью генной терапии дозу гена PTEN или генетически заингибировав его прямую мишень фосфоинозитид-3-киназу класса 1 (PI3K), что эквивалентно активации PTEN, можно продлить жизнь мышей .
| Активность | |
|---|---|
| Регуляция | |
| Классификация | |
| Типы |
|
| КФ 3.1.1: | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| КФ 3.1.2: | |||||||||||||||
| КФ 3.1.3: Фосфатазы | |||||||||||||||
| КФ 3.1.4: Фосфодиэстеразы | |||||||||||||||
| КФ 3.1.6: | |||||||||||||||
|
Нуклеазы (включая
дезоксирибонуклеазы и рибонуклеазы ) |
|
||||||||||||||