Ленточные черви
- 1 year ago
- 0
- 0

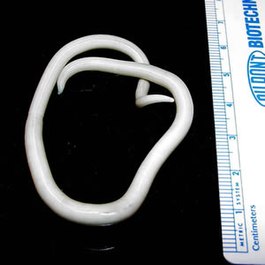
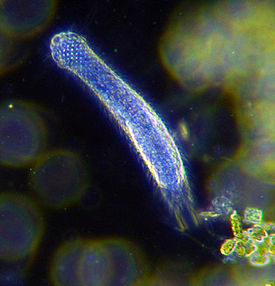
Многощети́нковые че́рви , или полихе́ты ( лат. Polychaeta , от греч. πολύς — много, греч. χαίτη — волос) , — класс кольчатых червей . В настоящее время этот класс насчитывает более 10 тысяч видов. В ископаемом состоянии известны с раннего кембрия . Наиболее известные представители: пескожил Arenicola marina и нереида Nereis virens .
Подавляющее большинство представителей — обитатели морских вод . Они заселяют океан от приливно-отливной полосы ( литорали ) до абиссали . Взрослые, как правило, — донные формы . Наибольшая глубина, на которой обнаружены многощетинковые черви — от 10 160 до 10 730 м. В Мировом океане полихеты заселили практически все биотопы дна и пелагиали — от интерстициали до гидротермальных оазисов океанических рифтовых зон. Очень немногие полихеты живут в пресных водоёмах (например, род в озере Байкал ), в лесной подстилке и в почве на глубине более 3 м (род и род ).
Большинство полихет ведет роющий образ жизни. Следующее место по численности занимают ползающие полихеты. Многие виды могут плавать, а некоторые даже ведут постоянно пелагический образ жизни . Наконец, имеются полихеты, которые формируют трубки и ведут в них сидячий образ жизни. Очень редки среди полихет паразиты, но некоторые виды паразитируют на других полихетах, морских звездах и на рыбах .
Этапом жизни личинок некоторых видов полихет (из семейств Amphinomidae , , , ) является пересечение с помощью течений Атлантического океана от Африки до Америки . Они так и называются трансокеаническими личинками. Многим из них удаётся за 2-3 месяца переплыть океан, затем осесть на дно, превратиться во взрослых червей и жить в прибрежном районе .

Полихеты характеризуются средним размером около 10 см в длину и 0,5–1,0 см в ширину. Интерстициальные виды обычно мелки, их размер не превышает 1 мм. С другой стороны, среди полихет есть крупные виды, достигающие метра в длину. Наиболее крупная полихета Eunice aphroditois 3 м длиной. Тело состоит из множества (иногда до нескольких сотен) колец-сегментов, в каждом из которых повторяется комплекс внутренних органов: парные целомические мешки , ассоциированные с ним половые протоки и органы выделения.
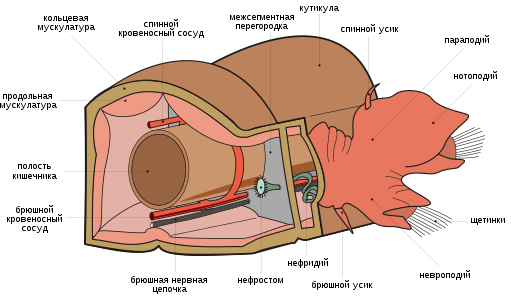
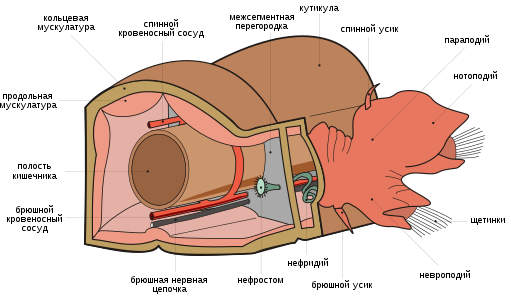
Отличительным признаком являются параподии — отходящие от каждого сегмента тела лопастевидные придатки, несущие хитиновые щетинки (хеты). У некоторых видов функцию жабр выполняет венчик щупалец на головном участке. Имеются глаза, иногда сложно устроенные, и органы равновесия ( статоцисты ).
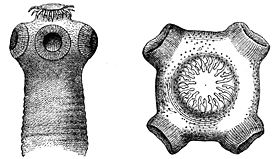
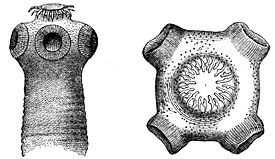
Простомиум (головная лопасть) — самый первый сегмент (счет сегментов тела начинается с перистомиума), всегда один, хотя и может вторично сегментироваться. На практике при определении границ простомиума удобно использовать положение ротового отверстия: простомиум — это сегмент, расположенный над или перед ртом, нередко образующий верхнюю губу. Простомиум может нести придатки. Если эти придатки примерно одинакового размера и формы, то их называют антеннами (головными щупальцами). Если же среди придатков резко выделяется одна пара, то эта пара получает название пальпы (щупики). У некоторых таксонов пальпы могут сливаться ( ), или разделяться на две пары (некоторые Onuphidae ), или образовывать жаберно-ловчий аппарат ( ), или ротовые щупальца ( ), или же смещаться на перистомиум ( ). Как антенны, так и пальпы могут иметь подставки, называемые, соответственно, церато- и пальпофоры, их дистальные части получают название церато- и пальпостили. Кроме того, на простомиуме могут быть разнообразно устроенные глаза: от простых пигментных пятен, до сложно устроенных оптических систем. Глаза могут располагаться как непосредственно на поверхности простомиума, так и на особых выростах — омматофорах. Форма простомиума часто имеет таксономическое значение.
Среди туловищных сегментов различают перистомиальный сегмент (сегменты) и собственно сегменты тела. Перистомиальный сегмент — перистомиум — следует непосредственно за простомиумом, с перистомиума начинается счет сегментов тела. Он несет с брюшной стороны ротовое отверстие, но в образовании ротового отверстия могут принимать участие и следующие сегменты (у Amphinomidae и ). Перистомиум, а также следующие за ним сегменты могут резко отличаться от расположенных за ними. Обычно в последнем случае следующие за перистомиумом сегменты весьма сходны с ним. В этом случае они получают название перистомиальных сегментов. Перистомиум может частично или полностью сливаться с простомиумом и/или следующими сегментами. Часто это слияние столь полно, что в некоторых случаях все ещё нет единого мнения относительно исходного количества сегментов и их границ. Слившиеся перистомиум и перистомиальные сегменты для краткости часто именуются перистомиумом. Часто перистомиум несет перистомиальные (щупальцевидные) усики, сходные по строению с антеннами, но в отличие от них перистомиальные усики гомологичны придаткам параподий. Количество пар перистомиальных усиков примерно соответствует количеству сегментов, образовавших перистомиум (1-2 пары па сегмент). Расположенное на перистомиуме ротовое отверстие ведет обычно в более или менее выворачивающуюся или выпячивающуюся глотку. Глотка у многих семейств вооружена хитиновыми челюстями (которые могут обызвествляться), а ее поверхность может быть покрыта мягкими кожистыми папиллами и/или хитиновыми зубчиками (парагнатами, шевронами и др.).
Пигидий (пигидиум) имеет обычно конусообразную форму с располагающимся на заднем конце апикально или смещенным несколько дорсально анальным отверстием. Как правило, пигидий несет усикообразные чувствительные придатки — уриты. Их число и размеры колеблются в широких пределах. Часто это пара хорошо развитых антенн. У некоторых ( и др.) на пигидии могут развиваться присасывательные приспособления (папиллы, лопасти и т. п.). На пигидии Sabellidae нередко имеются глазки.
Сегменты тела, как правило, несут по бокам параподии — более или менее выраженные выросты тела, снабженные щетинками . Границы между сегментами часто плохо выражены. На каждом сегменте может быть только одна пара параподий. Каждая параподия может иметь одну или две снабженных щетинками ветви: спинную — нотоподию и брюшную — невроподию. Параподии используются червями при ползании по дну водоема, а при плавании животного играют роль плавников. У червей, роющихся в грунте или живущих в трубчатых домиках, параподии полностью или частично редуцированы.
Покровы полихет состоят из однослойного кутикулярного эпителия и практически полностью лишены мерцательных клеток. Последние сохраняются лишь у личинок, где образуют мерцательные обручи, охватывающие головную лопасть и далее каждую метамеру. Ресничная локомоция существует на этой стадии онтогенеза до перехода к движению с помощью параподий. Сохраняется ресничный способ локомоции и соответственно ресничные обручи у взрослых особей тех полихет, которые считаются вторично измельчавшими формами неотенического происхождения ( Ophryotrocha ). Наконец, мерцательный эпителий сохраняется у некоторых форм не для локомоции, а для создания токов воды по телу, необходимых при некоторых условиях жизни. В частности, поэтому мерцательные клетки сохраняются на жабрах у ряда полихет. Поля мерцательных клеток находят и у трубкоживущих полихет, у которых они могут, например, нести к ротовому отверстию частички пищи. Или, напротив, выносить из трубки фекалии.
Кутикула полихет плотная и одновременно гибкая, не имеющая такого мощного, сложно слоистого строения, как у нематод . Её структура варьируется у разных полихет, представляя собой или толстый единый слой из полисахаридов и белков- альбуминов , или же решетку коллагеновых фибрилл, чередующихся с гранулярными слоями. В таком случае ультрамикроскопические исследования обнаруживают в кутикуле два слоя: эпи- и базальную кутикулу. Первая состоит из мелкогранулярного или тонковолокнистого матрикса и электронно-плотного слоя, в котором располагаются своеобразные сферические тела. Здесь обнаружены кислые мукополисахариды . Базальная кутикула состоит из фибрилл полисахаридной или гликопротеидной природы, образующих мощную беспорядочную или, наоборот, упорядоченную решетку коллагеновых волокон.
Во всех случаях кутикула не сплошная. Эпидермальные клетки образуют многочисленные ультрамикроскопические выпячивания — микровилли, которые проникают сквозь поры решетки фибрилл и отчасти даже возвышаются над поверхностью кутикулы. Благодаря этому осуществляется свободный обмен веществ между эпидермисом и окружающей средой.
Эпидермис полихет содержит большое количество железистых клеток. Они могут открываться на поверхности кутикулы. Среди железистых клеток преобладают слизистые, которые вырабатывают слизь для защиты тела (например, у Anaitides mucosa ). Эпителий подостлан слоем ткани основного вещества в виде тонкой пограничной пластинки с сильно развитыми преколлагеновыми волокнами и обычно лишенной клеток.
Покровы червей, ведущих активную жизнь на дне водоема, отличаются хорошо развитой кутикулой. Напротив, у червей, плавающих в толще воды, зарывающихся в грунт или строящих трубчатые домики, кутикула очень тонкая. Выделения железистых клеток служат строительным и цементирующим материалом при постройке трубок, в которых живут некоторые многощетинковые черви.
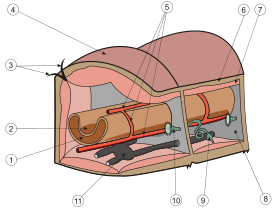
Мускулатура полихет лежит непосредственно под пограничной пластинкой. Она представлена расположенным снаружи слоем кольцевых мышц и, глубже, слоем продольных мышц. Имеется и дорсовентральная мускулатура, у полихет теснейшим образом связанная с развитием стенок диссепиментов и мезентериев целома. Существуют также мышечные пучки, относящиеся к системе локомоторных органов (параподиев) и специальные мышечные образования различных органов (кишечника, кровеносных сосудов и т. п.).
Пищеварительный тракт полихет состоит из трех отделов: переднего эктодермального (стомодеального) — глоточного; среднего ( энтодермального ) — усваивающего; заднего (проктодеального) — выводящего. Стомодеальный отдел начинается поперечным ротовым отверстием, за которым следуют ротовая полость, мускулистая глотка (фаринкс) и эзофагус («пищевод»). У многих хищных червей в глотке есть хитиновые зубцы, служащие для схватывания добычи . Полихеты демонстрируют большое разнообразие в строении стомодеального отдела. Особенно это касается фаринкса. Это обусловлено тем, что полихеты освоили самые разнообразные биотопы и очень разные стратегии добывания пищи. Желудок имеется только у сидячих полихет. У нереисов имеются крупные слепые отростки пищевода, которые выделяют пищеварительные ферменты. Энтодермальная средняя кишка, в отличие от переднего отдела кишечника, довольно подвижного вдоль продольной оси тела, прочно покоится, закрепленная диссепиментами и охваченная вдоль всей своей длины мезентериями. Она обычно прямая, без извивов, что является следствием её зафиксированности стенками целомических мешков. Транспорт пищевых частиц в неподвижном кишечнике осуществляется главным образом с помощью ресничек. Для этого, как правило, вдоль вентральной поверхности средней кишки тянется продольная бороздка или валик, несущие мерцательные клетки. В средней кишке происходит переваривание пищи и усвоение питательных веществ. Короткая эктодермальная задняя кишка открывается анальным отверстием терминально. Однако порошица может смещаться дорсально или вентрально.
Для видов, живущих в трубках, характерны разные формы удаления экскрементов, обеспечивающие вывод этих структур из трубки наружу. Наиболее оригинален способ, характерный для представителей семейства . Эти виды живут в вертикально расположенных трубках, и передним концом они обращены вверх. Задняя часть тела этих полихет сильно редуцирована, от нее осталась лишь выводящая фекалии трубка, лежащая в задней части спинной стороны животного. От конца этой трубки по спинной стороне тела к переднему концу (вверх) тянется ресничный желобок, по которому плотные фекалии и удаляются из трубки.
Дыхание у мелких форм осуществляется поверхностью эпидермиса. У более крупных видов в определенных местах эпидермиса образуются участки, снабженные сетью сосудов и связанной с ней мощной капиллярной сетью. У части видов возникают особые тонкостенные выросты тела или параподий, которые являются специализированными органами дыхания, такие образования традиционно называют жабрами. У ползающих форм в жабры, если они имеются, чаще всего превращаются спинные усики параподий. Необходимость в жабрах возникла в результате сильного уменьшения количества диффундирующего через покровы растворенного в воде кислорода, которое является следствием сокращения относительной поверхности тела в связи с увеличением его линейных размеров и развития кутикулярных покровов. Форма жабр сильно изменчива: встречаются нитевидные (в наиболее простом случае), пузыреобразные, гребне- и древовидные. У ряда полихет в них заходят отроги целомических мешков, у других, кроме этого, жабры снабжены сетью капилляров кровеносной системы.
Транспортные функции полихет осуществляются циркуляцией жидкости в целоме и в кровеносной системе. Если диссепименты полностью утрачиваются (что характерно для представителей семейства Glyceridae ), то утрачивается и кровеносная система, транспортные функции тогда осуществляются только целомом. У части мелких полихет кровеносная система также может отсутствовать.
Строение кровеносной системы полихет соответствует общей схеме кровеносной системы аннелид . Она замкнутая, включает два главных продольных сосуда — пульсирующий спинной и несокращающийся брюшной. Метамерные сосуды представлены только поперечными, которые не составляют полного кольца (как в случае с кольцевыми метамерными сосудами), они от спинного и брюшного магистральных сосудов направляются к стенке тела, параподиям или каким-либо другим органам, где разбиваются на систему капилляров (после объединения капилляров в более крупные сосуды кровь по ним возвращается в какой-либо из продольных сосудов). От брюшного сосуда отходят два поперечных, один из которых несет кровь к спинной ветви параподии, другой — к брюшной. Эти же сосуды снабжают кровью стенку тела и жабры, если они имеются. Кровь от параподий и стенки тела поступает в спинной сосуд. Оттуда кровь идет к кишечному плексусу, от кишечного плексуса — в брюшной сосуд.
Дыхательные пигменты могут быть представлены гемоглобином , что является наиболее частым вариантом, гемэритрином и хлорокруорином . Гемоглобин и хлорокруорин находятся в плазме крови, гемэритрин, наличие которого характерно для одного семейства полихет, содержится в клетках крови.
Как общее правило, в каждом туловищном сегменте полихет имеется пара протонефридиев или пара метанефридиев . Нефридии характеризуются эктодермальным про-исхождением. Внутренний конец каждого нефридия лежит в целоме, возле его задней стенки. Канал нафридия прободает диссепимент и в следующем сегменте открывается наружу отверстием — нефропором — на боковой стороне тела. Протонефридии характерны для мелких видов полихет, не имеющих кровеносной системы. Крупные виды полихет, как правило, имеют метанефридии. Последние представляют собой извитые трубочки, начинающиеся в полости тела воронкой с ресничками. Внешний конец метанефридиев открывается либо непосредственно наружу, либо в продольный общий выделительный канал. Конечный отдел метанефридия имеет расширение — мочевой пузырь.
Нервная система представлена надглоточным ганглием (головным мозгом) и брюшной нервной лестницей или брюшной нервной цепочкой (брюшной мозг). У форм с хорошо развитыми органами чувств надглоточный ганглий может приобретать крупные размеры и быть разделенным на несколько долей. Головной мозг играет роль самостоятельного центра, который регулирует деятельность всей нервной системы туловища. Как норма, он располагается в головной лопасти. Дорсально и с боков он обычно не отделен от покровного эпителия, тогда как снизу подстелен субэпителиальной пограничной пластинкой. Объем мозга сильно варьируется: у эррантных крупных Aphroditidae суммарное количество клеток мозга доходит до 14 тысяч клеток, а у Polydora ciliata — седентарной формы — едва достигает 800. В своем наиболее развитом виде брюшной мозг представляет собой пару мощных нервных стволов, идущих вентрально вдоль всей длины тела параллельно друг другу. Как правило, они сближены между собой вдоль медиальной линии вплоть до слияния в единое анатомическое целое, но у , например, они довольно заметно удалены друг от друга и благодаря комиссурам (поперечным нервам) образуют брюшную нервную лестницу. Заметно варьируется и глубина залегания брюшного мозга. У некоторых полихет он лежит субэпителиально ( , и др.). Прослеживаются все стадии погружения брюшного мозга из-под покрова в стенку тела, под кольцевую мускулатуру и далее в полость целома у ряда семейств полихет. Типичным для полихет является наличие педальных ганглиев, расположенных у основания параподий.
Щупальца и пальпы несут на своей поверхности механорецепторы (осязательные) и хеморецепторы. Перистомиальные усики, как щупальца и пальпы, снабжены механорецепторами и хеморецепторами. В задней части простомиума находятся светочувствительные органы. Эти органы представлены в разном числе и могут быть простыми глазками, состоящими всего из двух клеток, или достаточно сложными глазами, включающими тысячи чувствующих клеток, а также линзы. Более сложно устроенные глаза встречаются у плавающих видов. Кроме простомиума, глаза могут располагаться на сегментах тела, на пигидиуме, на жабрах. На простомиуме располагаются нюхальные органы, или обонятельные ямки — небольшие углубления покровов, на дне которых расположены ресничные клетки, выполняющие хеморецепторную функцию.
Большинство многощетинковых червей раздельнополы. В каждом сегменте находится пара гонад. Они залегают в соединительной ткани и связаны с септами, кровеносными сосудами и целомической выстилкой. Гонады часто присутствуют во всех сегментах, однако у некоторых полихет они приурочены к особым генитальным сегментам. Гермафродитные полихеты немногочисленны. Среди них встречаются виды (некоторые Sabellidae ), у которых передние брюшные сегменты производят яйца, а задние — сперму. Как правило, гаметы выводятся во внешнюю среду через метанефридии или через разрывы стенки тела.
Особенности питания разных полихет тесно связаны с образом их жизни. Многие роющие формы и сидячие обитатели норок и трубок — грунтоеды , использующие содержащуюся в грунте органику. Одни из них заглатывают грунт непосредственно ртом или с помощью снабженной бульбусом глотки, которая способна выпячиваться наружу и характеризуется слабым развитием мышц. Это так называемые заглатывающие детритофаги , к числу которых относятся, например, пескожилы ( ), мальданиды ( ) и напоминающие дождевых червей полихеты из групп Orbiniidae и Capitellidae. В противовес этому собирающие детритофаги , например представители и , сначала собирают органические частицы специализированными придатками, которые затем отправляют пищу в рот. Представители Terebellidae собирают пищу с помощью способных растягиваться и сокращаться простомиальных щупалец, а спиониды (Spionidae) делают это длинными перистомиальными пальпами.
Плотоядные и растительноядные формы и полихеты-падальщики обычно представлены подвижными (бродячими, или эррантными) формами, которые ползают по субстрату или ловят планктон ( ), но некоторые из них живут в трубках или активно роются в грунте. Для захвата добычи или падали эти полихеты имеют хорошо развитую, мускулистую, способную выворачиваться глотку. Вывернутая глотка может иметь форму мускулистой трубки и служит для захвата добычи ( Phyllodocidae ). Иногда она снабжена толстой, похожей на терку нижней губой ( Amphinomidae ) или несет челюсти с хватательными ( Nereidae , Eunicidae , ) или ядовитыми ( Glyceridae ) зубами. Роговые челюсти построены из задубленного белка. Как только пища обнаружена, глотка мгновенно выворачивается. Расположенные на её конце челюсти при этом выносятся вперед и раскрываются. Челюсти захватывают пищу, после чего глотка столь же быстро втягивается. Хотя в ряде случаев полихеты имеют специальные мышцы-протракторы, выворачивание глотки, как правило, происходит за счет увеличения давления целомической жидкости в результате сокращения мышц стенки тела. Когда эти мышцы расслабляются и давление падает, глотка втягивается мышцами-ретракторами.
Все полихеты, питающиеся взвесью (сестонофаги), — сидячие животные, обитающие в трубках. Последние могут быть погружены в грунт или прикреплены к раковинам моллюсков, скалам и другим твердым поверхностям. Для улавливания взвешенных в воде частиц такие полихеты используют специальные придатки, несущие реснички и обладающие большой площадью поверхности (у Sabellidae , например, это перистая «корона») или выделяют слизистую сеть, через которую активно прогоняется и фильтруется вода ( Chaetopterus ).
Очень небольшое число видов полихет ведет паразитический образ жизни. ( ) поселяется в целоме других полихет и может достигать почти таких же размеров, что и хозяин. Некоторые мизостомиды паразитируют в морских звездах . К числу эктопаразитов относятся питающиеся кровью , которые прикрепляются к плавникам некоторых морских рыб.
Большинство полихет легко восстанавливает утраченные части тела. В общем, способность к регенерации лучше выражена у гомономных форм. Экспериментальные исследования показали, что у некоторых видов возможно полное восстановление тела всего из одного сегмента. Регенерация идет под контролем нервной и эндокринной систем.
Полихеты реализуют различные способы движения. Роющие полихеты передвигаются в грунте за счет перистальтики. У некоторых роющих полихет в движении принимает участие глотка, которая, выворачиваясь вперед, заякоривается в грунте, затем червь сокращает глотку и подтягивается вперед. Иногда движение подобного рода осуществляется только за счет работы глотки. При ползании параподии и щетинки работают, как ноги, поочередно отталкиваясь от субстрата во время рабочего удара и приподнимаясь над субстратом во время возвратного движения. Движения многочисленных параподий хорошо скоординированы. Параподии, расположенные на противоположных сторонах одного сегмента, движутся в противофазе. Движения параподий, расположенных на одной стороне тела, смещены относительно друг друга на долю фазы. В движении ползания принимает участие и ундуляция тела в латеральном направлении. Аналогичные движения используются полихетами и при плавании.
Оплодотворение у полихет наружное. Они откладывают яйца непосредственно в воду, где происходит их оплодотворение и из них развиваются пелагические личинки ( трохофоры ), более или менее долгое время существующие в толще воды и способные к активному питанию. Часть видов полихет, обитающих в грунте или в интерстициали, освоила внутреннее оплодотворение и, соответственно, обладает копулятивными органами . Есть виды, у которых сперма вводится в тело самки через кожу, за счет её прокалывания, в других случаях сперматофор приклеивается с помощью копулятивного органа к поверхности тела самки, сперматозоиды своими средствами пробуравливают кожу и поступают в целом самки. У части видов с внутренним оплодотворением наблюдается забота о потомстве, выражающаяся в формировании защитного кокона для кладки и даже его ношения на теле самки.
После некоторого периода планктонной жизни трохофора опускается на дно, где происходит метаморфоз . Переднее полушарие трохофоры формирует щупальца и пальпы, а также другие органы и постепенно превращается в простомиум. Околоротовой отдел трохофоры перестраивается в перистомиум. Заднее полушарие личинки сильно вытягивается и подразделяется сразу на несколько сегментов, которые называются ларвальными сегментами. Самый задний конец заднего полушария пребразуется в пигидиум. В каждом сегменте развивается пара целомических мешков. В процессе матаморфоза формируется головной мозг, на брюшной стороне в виде парного валика эктодермы закладываются брюшные нервные стволы. В дальнейшем они вступают в связь с головным мозгом при помощи окологлоточных коннектив. Из эктодермы развиваются и органы чувств — глаза, пальпы. Так из несегментированной, первичнополостной трохофоры формируется следующая личиночная стадия — метатрохофора, характеризующаяся сегментацией и метамерным целомом. Метатрохофора некоторое время плавает или ведет донный образ жизни, не меняясь существенно, а затем испытывает дальнейший метаморфоз. На переднем крае анальной лопасти образуется зона роста, клетки которой непрерывно размножаются. Область, лежащая впереди неё, состоит из быстро растущих еще не дифференцированных тканей. В этой зоне формируются новые сегменты (постларвальные) и последовательно один за другим отделаются по направлению кпереди. Процесс продолжается до тех пор, пока не образуется столько сегментов, сколько их у взрослого червя. Следовательно, тело взрослого животного состоит из различных по происхождению отделов: головной лопасти или простомиума, представляющей собой видоизмененное переднее полушарие трохофоры; нескольких ларвальных сегментов; многочисленных постларвальных сегментов и анальной лопасти или пигидиума, происходящей из самого заднего участка трохофоры.
Из мезодермы формируются мускулатура кожно-мускульного мешка и кишечника, выстилка целома, гонады и целомодукты, из эктодермы — нервная система, каналы метанефридиев, передняя и задняя кишка, из энтодермы — средняя кишка.
Некоторые виды способны размножаться бесполым путём. Выделяют два варианта бесполого размножения: и . В случае архетомии тело червя сперва разделяется на фрагменты, а после достраивает передний и задний концы тела. Паратомия же подразумевает обратную последовательность событий: в ходе этого процесса образуется цепочка сцепленных друг с другом разноимёнными концами тела червей.
Половой диморфизм, как правило, не выражен. Наиболее известные случаи такового — эпитокные стадии полихет из семейств Nereidae и . Половой диморфизм возникает лишь у половозрелых особей, готовящихся к нересту. В этом случае передняя (атокная) часть тела полихеты, лишенная гонад, отличается и по строению параподий, которые, как правило, развиты слабее, и по окраске тела. Последнее обусловлено цветом половых продуктов, просвечивающих сквозь полупрозрачные покровы тела. Это приводит к появлению полового диморфизма, так как половые продукты самцов и самок окрашены по-разному. Задняя часть тела, кроме окраски, характеризуется большей толщиной, сильнее развитыми параподиями и называется эпитокной.
Процесс разможения полихет контролируется гормонами, которые имеют нейросекреторную природу. Их продуцируют клетки мозга или (у представителей ) нервные элементы специализированной передней кишки, приспособленной для сосания. У полихет, которые размножаются один раз и затем погибают (например, Nereidae и Syllidae), все этапы полового размножения находятся под гормональным контролем. Это относится и к продукции гамет, и к формированию эпитокной формы. У видов, размножающихся несколько раз, с помощью гормонов преимущественно регулируется развитие гамет, в первую очередь яиц. Влияние гормонов в основном распространяется лишь на эти процессы.
Крайне своеобразный феномен эпитокии проявляется в период размножения многих полихет. Особенно хорошо это явление изучено у представителей Nereidae, Syllidae и Eunicidae. Эпитокия — это образование пелагической эпитокной особи, способной к половому размножению, из бентосной атокной особи, не способной производить половые продукты. Эпитокия выражается не только в достижении половой зрелости и производстве гамет, но и в перестройке ряда структур, непосредственно с репродукцией не связанных. Эти изменения направлены на усиление способности червей активно плавать и обнаруживать возможного партнера. Чаще всего у эпитокных форм увеличиваются глаза, модифицируются параподии и щетинки, увеличиваются размеры сегментов и более мощного развития достигают мышцы. Атокная особь может давать начало одной или нескольким эпитокным. В первом случае исходная атокная особь сама превращается в эпитокного червя, как это имеет место у представителей Nereidae. Во втором случае дифференцировке подвергается только задняя половина тела, которая позднее отпочковывается и становится самостоятельной эпитокной особью. Содержащие гаметы сегменты эпитокной особи часто очень сильно видоизменяются, так что тело червя оказывается четко разделенным на два отдела. Например, эпитокные формы Nereis irrorata и N. succinea имеют большие глаза и редуцированные простомиальные пальпы и щупальца. Передние 15—20 сегментов туловища изменены незначительно, зато следующие за ними сегменты, формирующие эпитокную область и набитые гаметами, увеличены; их параподии несут «веера» длинных, лопатовидных, плавательных щетинок. У палоло передний конец червя не изменен, в то время как эпитокная область представляет собой цепочку набитых яйцами сегментов.
Ресничные поля у личинок полихет могут быть двух типов: ресничные поля, приспособленные для поступательного движения, и ресничные поля тактильного свойства. К ресничным полям, осуществляющим функцию осязания, относятся апикальный султан ресничек, а также акротрох и тактильные реснички, расположенные на эписфере или простомиуме, на анальной лопасти или в составе ресничных колец. Ресничные поля, осуществляющие функцию движения, можно разделить на собственно кинетические и гидрокинетические. Первые осуществляют движение самой личинки. Это прото-, мезо-, тело-, ното- и гастротрохи. Вторые служат для привлечения новых масс воды к телу и дистанционным органам чувств, для доставки кислорода, удаления отторгнутых отбросов и получения информации о запахах. Это реснички в области затылочных органов, в области ротового поля, ресничной ямки и невротрохоид.
У пелагических личинок всегда развита одна или несколько пар глаз. Некоторые виды в развитии утрачивают глаза. Обычно это роющие формы. У других провизорные глаза заменяются дефинитивными. Чаще всего глаза, возникающие у трохофоры, продолжают служить и во взрослом состоянии. Глаза могут быть простыми и сложными. Сложные глаза собраны группами на простомиуме или на жабрах. Одиночные, или простые глаза, как и сложные, могут быть с хрусталиками и без них.
У личинок полихет развита пигментация двух типов. Во-первых, пигменты, тонко распыленные внутри клеток тела. Во-вторых, пигменты в виде меланофоров, хроматофоров и зерен меланина или пигментов другого свойства в эпидермисе. Часто на ранних стадиях пигмент развит слабее, а на более поздних очень интенсивно. Однако при переходе к существованию на дне он исчезает или заменяется другим.
Личинки, которые ведут длительный пелагический образ жизни, снабжены специальными провизорными щетинками. Они увеличивают площадь опоры и уменьшают остаточный вес. Например, личинки семейства — митрарии — снабжены двумя длинными пучками провизорных щетинок, которые сбрасываются в момент катастрофического метаморфоза .
Наряду с провизорными у личинок одновременно могут быть развиты и дефинитивные щетинки. Они часто располагаются в ларвальных сегментах, которые у взрослых порой не должны нести щетинок. Однако специализированные дефинитивные щетинки закладываются очень рано и могут служить хорошим признаком для идентификации вида. Например, специализированные щетинки 5-го сегмента у Polydora или специализированные щетинки торакального отдела Chaetopterus .
В процессе метаморфоза полихет резко меняется весь план строения. Несегментированное тело становится сегментированным. Ортогональная нервная система трохофоры превращается в сложно дифференцированную дефинитивную нервную систему. Первичная полость тела замещается вторичной. Протонефридии заменяются метанефридиями . Появляются гонады .
Полихеты имеют очень различно развитую нервную систему. Чем сильнее дифференцирована нервная система во взрослом состоянии, тем раньше появляются в процессе развития соответствующие закладки. Например, стебельчатые тела головного мозга закладываются уже у метатрохофоры Aphroditidae . В противоположном случае у личинок семейства головной мозг оказывается более дифференцированным, чем у взрослых форм.
Кожно-мускульный мешок личинок полихет состоит из покровного, железистого и ресничного эпителия, кольцевой и продольной мускулатуры. Кольцевая мускулатура развита заметно слабее продольной. Продольная мускулатура расположена или четырьмя продольными лентами (чаще у ) либо сплошным тонким слоем (чаще у ).
Первичная полость тела по объему очень различна у разных личинок полихет. Объем полости находится в обратной зависимости от скорости движения и в прямой зависимости от продолжительности пребывания личинки в планктоне .
Метамерия целома в ларвальном теле, как правило, отсутствует. Парные протонефридиальные органы выделения личинок меняются на органы выделения метанефридиальные или смешанные.
Кишечник личинок состоит из трех отделов. Передний отдел включает пищевод и парные карманы зачатка дефинитивной глотки. Средний отдел — энтодермальная кишка — всегда состоит из двух камер. Задний отдел — прямая кишка.
Кровеносная система развита не у всех полихет, но там, где она есть, всегда связана с появлением постларвального тела.