

Молекулярный человек (Marvel Comics)
- 1 year ago
- 0
- 0


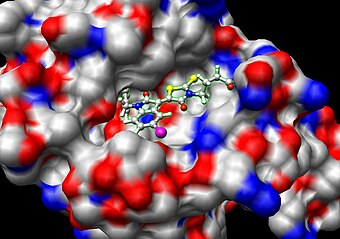
Молекуля́рный до́кинг ( англ. Molecular docking ) — метод молекулярного моделирования , позволяющий предсказать наиболее выгодную для образования устойчивого комплекса ориентацию и конформацию одной молекулы ( лиганда ) в сайте связывания другой ( рецептора ) . Данные о положении и конформации партнеров используются для предсказания силы взаимодействия посредством так называемых оценочных функций. В случае, если лиганд является макромолекулой , докинг называют макромолекулярным .
Молекулярный докинг можно представлять как поиск оптимального положения «ключа» (лиганда) в «замке» (рецепторе) . В данном случае молекулы рассматриваются как жёсткие тела. Однако в реальности в процессе докинга лиганд и белок изменяют конформации для достижения наилучшего связывания. Изменения конформации белка могут включать движения и доменов . Такой процесс, ведущий к успешному связыванию, называют «индуцированным соответствием» .
Молекулярный докинг используется для моделирования процесса молекулярного узнавания. Обычно необходимо найти оптимальную конформацию лиганда. Данное положение достигается в случае, когда свободная энергия связывания минимальна. .
Комплексы таких биологически значимых молекул, как белки, нуклеиновые кислоты , углеводы и липиды , играют ключевую роль в передаче химического сигнала. К тому же, относительная ориентация двух взаимодействующих молекул может влиять на тип произведённого сигнала (будет он ингибирующим или каталитическим ). Поэтому взаимодействие между биологическими молекулами важно для предсказания как типа, так и силы производимого сигнала .
Докинг часто используется для предсказания аффинности и активности небольшой молекулы лекарства по отношению к белку-мишени. Таким образом, молекулярный докинг, являясь одним из этапов при разработке , играет важную роль в данном процессе .
Одним из преимуществ молекулярного докинга является возможность его автоматизации. В рамках задачи разработки препарата появляется возможность скрининга . Молекулярный докинг позволяет определить наиболее оптимально взаимодействующее соединение — лекарство из ряда близких по составу аналогов .
Одним из методов, применяемых при разработке лекарственных препаратов, является . Метод основывается на поиске небольших фрагментов, обладающих невысокой аффинностью связывания с мишенью, и их дальнейшего комбинирования с целью поиска соединения с высокой аффинностью. Фрагментный дизайн применяется для поиска сильнодействующих ингибиторов. Подобная задача решается с помощью различных методов. К ним относятся некоторые виды ЯМР-спектроскопии , , метод микроскопического термофореза , плазмонный резонанс и другие . Молекулярный докинг, в свою очередь, также позволяет решить подобную задачу путём сканирования библиотек различных соединений, как низкомолекулярных, так и комплексных, и оценивать их аффинность .
Докинг может быть использован в биоремедиации для поиска загрязнителей окружающей среды, разлагаемых некоторыми ферментами .
Однако бывают случаи, когда неизвестно непосредственно само место взаимодействия. Тогда применим так называемый «слепой» докинг . Различные вариации данного подхода реализованы в алгоритмах: MolDock , Fragment Hotspot Maps , DoGSiteScorer .
Среди фундаментальных направлений применения молекулярного докинга можно выделить :
Существуют различные подходы при моделировании докинга. Один из подходов использует технику соответствия, которая описывает белок и лиганд как дополнительные поверхности . Другой подход моделирует фактический процесс докинга, в котором вычисляются попарные энергии взаимодействия . У обоих подходов есть существенные преимущества, а также некоторые ограничения .
«Жёстким» называется докинг, при котором длины связей, углы и торсионные углы партнеров докинга остаются неизменными в процессе моделирования. Однако в результате взаимодействия с другим белком или лигандом происходят конформационные изменения как самого остова белка, так и боковых цепей. Подвижность остова, в свою очередь, может быть разделена на два типа: подвижность больших участков белка — доменов, так называемое движение «сдвига», и подвижность отдельных частей, таких как петли. В данном случае «жесткий» докинг некорректно описывает взаимодействия. Поэтому существуют некоторые дополнительные алгоритмы «гибкого» докинга. Они допускают конформационные изменения, в результате чего данный подход позволяет получать оценки взаимодействий наиболее приближенные к естественным. Однако подсчёт всех возможных конформационных изменений с учётом движения на данном уровне развития компьютеров занял бы огромное время. Более того, большое количество степеней свободы также может приводить к увеличению количества ложно положительных результатов. В связи с данными проблемами, возникает необходимость рационально выбирать небольшое подмножество возможных конформационных изменений для проведения моделирования .
«Гибкий» докинг также может быть использован в рамках докинга низкомолекулярного соединения. Однако в данном случае разрешается вращение вокруг каких-либо связей в молекуле самого лиганда, белок при этом остается «жёсткой» структурой
Докинг также можно разделить на однократный ( англ. single ) и последовательный ( англ. sequential ) . Последовательный докинг применяется, в основном, для докинга нескольких низкомолекулярных соединений (лигандов). После докинга одного из лигандов в отдельный файл сохраняется структура белка с данным лигандом. Далее алгоритм повторяется, и реализуется докинг для второго лиганда в ранее сохраненную структуру. Данный подход может быть полезен при поиске аллостерических центров .
Геометрическое соответствие (методы определения взаимозависимости формы) описывается для белка и лиганда как ряд особенностей, которые определяют их оптимальное взаимодействие . Эти особенности могут включать как саму , так и описание дополнительных особенностей поверхности. В этом случае молекулярная поверхность рецептора описывается с точки зрения её доступности для растворителя , а молекулярная поверхность лиганда описывается с точки зрения её соответствия описанию поверхности рецептора. Взаимозависимость между двумя поверхностями составляет описание соответствия формы, которое может помочь обнаружить различные положения лиганда. В другом подходе нужно описать гидрофобные особенности белка, используя повороты в атомах главной цепи. Ещё один подход может быть основан на преобразовании Фурье .
В этом подходе белок и лиганд отделены некоторым физическим расстоянием, и лиганд находит своё положение в активный центр белка после определённого числа «шагов». Шаги включают преобразования твёрдого тела , такие как перемещение и вращение , а также внутренние изменения структуры лиганда, включая угловые вращения. Каждый из этих шагов в пространстве изменяет полную энергетическую оценку системы, и, следовательно, она вычисляется после каждого движения. Очевидное преимущество этого метода состоит в том, что это позволяет исследовать гибкость лиганда во время моделирования, тогда как методы взаимозависимости формы должны использовать некоторые другие подходы, чтобы узнавать о подвижности лиганда. Другое преимущество состоит в том, что процесс физически ближе к тому, что происходит в действительности, когда белок и лиганд приближаются к друг другу после молекулярного распознавания. Неудобство этой техники — то, что она занимает время, чтобы оценить оптимальное решение докинга, так как необходимо исследовать довольно большой .
Первое, что необходимо для проведения скрининга молекул докингом — это структура интересующего белка. Обычно структура определяется биофизическими методами ( рентгеноструктурный анализ или ЯМР-спектроскопия ), также она может быть получена . Структура белка вместе с базой данных потенциальных лигандов служат входом для программы докинга. Успех докинга зависит от двух компонентов: алгоритма поиска и оценочной функции .
Успешный докинг требует выполнения двух условий :
Для многих случаях, к примеру, для антител и конкурентных ингибиторов сайт связывания известен. В других случаях сайт связывания может быть определён по данным мутагенеза или филогении . Конфигурации, в которых атомы белков перекрываются (так называемый клеш, от англ. clash ) всегда исключаются .
После отсеивания комплексов с клешами измеряется энергия каждой структуры (модели комплекса) с помощью так называемой скоровой (оценочной) функции. Последняя должна различить «достоверную» структуру выше как минимум 100000 альтернатив. Это сложная вычислительная задача, поэтому было разработано множество методов её решения. Алгоритмы можно разделить на детерминированные и стохастические .
С математической точки зрения докинг представляет собой поиск глобального минимума функции , заданной на многомерном пространстве всех возможных способов связывания лиганда с белком. Пространство поиска в теории состоит из всех возможных положений в пространстве и конформаций белка, связанного с лигандом. Однако на практике, при имеющихся вычислительных ресурсах, невозможно полноценно исследовать пространство поиска — это бы потребовало вычисление всех возможных сдвигов каждой молекулы (молекулы динамичны и существуют как ансамбль конформационных состояний) и всех ротационных и позиционных положений лиганда относительно белка при заданном уровне детализации. Большинство программ докинга учитывают все конформационное пространство вариантов для лиганда («подвижный» лиганд), и некоторые также пытаются моделировать «подвижный» белок-рецептор. Каждая фиксированная позиция этой пары в пространстве называется решением докинга .
Алгоритмы поиска наилучшего связывания могут быть разделены на следующие категории: систематические методы, случайные или стохастические эвристические методы, методы молекулярной динамики и термодинамические методы .
Методами, гарантирующими нахождение глобального минимума за конечное число шагов, являются систематические методы, то есть методы последовательного перебора всех возможных положений лиганда в активном центре белка-мишени. Однако ввиду большого количества требуемых вычислений этот метод требует введения значительных упрощений. Широко распространены другие методы глобальной оптимизации, которые не гарантируют нахождение глобального минимума за конечное число шагов программы, однако на практике оказывается, что они способны отыскивать такие минимумы гораздо быстрее, чем методы систематического перебора. Подобные методы можно разделить на две большие группы: эвристические и термодинамические .
Эвристические методы используют некие эмпирические стратегии поиска глобального минимума, ускоряющие процедуру по сравнению с простым сканированием гиперповерхности. Наиболее известны и популярны следующие эвристические методы :
К термодинамическим методам относят моделирование отжига .
В методах типа Монте-Карло исходная конфигурация уточняется путем принятия или отвергания шагов (итеративных изменений некого набора параметров), в зависимости от значения оценочной функции (то есть скора структуры) (см. критерий Метрополиса ), пока не будет предпринято определённое количество шагов. Предполагается, что сходимость к наилучшей структуре будет происходить из большого класса начальных, только одну из которых необходимо учитывать. Исходные структуры могут быть гораздо быстрее проанализированы «грубыми» ( англ. coarsed ) методами. Трудно найти скоровую функцию, которая бы одновременно хорошо отличала «хорошую» структуру и сходилась ней с большого расстояния (в семплируемом пространстве). Поэтому было предложено использование двух уровней приближения («грубое» и «точное») с различными функциями оценки. Вращение может быть введено в Монте-Карло как дополнительный параметр для шага .
Методы Монте-Карло являются стохастическим и не гарантируют исчерпывающий поиск, следовательно, лучшая конфигурация может быть пропущена даже при использовании оценочной функции, которая в теории её отличает. Насколько серьёзно влияет эта проблема на результаты докинга, пока точно не установлено .
Данный подход реализован в алгоритме (англ.) . RossettaCommons . Дата обращения: 27 апреля 2020. .
В результате докинга генерируется большое количество потенциальных положений лигандов, некоторые из которых сразу отклоняются из-за наличия столкновений с молекулой белка. Остальные оцениваются с использованием функции оценки, которая принимает текущее решение докинга в качестве входных данных и возвращает число, указывающее вероятность того, что решение докинга представляет благоприятное связывающее взаимодействие. Таким образом, может быть оценена эффективность связывания одного лиганда относительно другого .
В современных алгоритмах докинга, можно выделить три основных типа оценочных функций: основанные на силовых полях, эмпирические и статистические.
Большинство скоринговых функций основаны на физике силовых полей молекулярной механики , которые оценивают энергию решения докинга в пределах сайта связывания. Различные вклады в энергию решения докинга можно записать в виде уравнения :
Компоненты уравнения включают в себя эффекты растворителя, конформационных изменений в белке и лиганде, свободной энергии из-за взаимодействий белок-лиганд, внутренних вращений, энергии ассоциации лиганда и рецептора с образованием единого комплекса и свободной энергии из-за изменений в колебательных режимах. Низкая (отрицательная) энергия указывает на стабильную систему и, следовательно, вероятное связующее взаимодействие .
Эмпирические оценочные функции, в отличие от основанных на силовых полях, включают компоненты, описывающие межмолекулярные контакты более простым способом. Прямых аналогий с парными межмолекулярными физическими взаимодействиями в данном случае нет. Предсказательная способность такого подхода сильно зависит как от самих составляющих, так и от коэффициентов, с которыми они входят в уравнение. Межмолекулярные взаимодействия представлены в виде линейной комбинации слагаемых, описывающих различные виды контактов: водородные связи, гидрофобные взаимодействия, взаимодействия с ионами металлов и другие. Упрощение, например для координационных связей с ионами металлов или гидрофобных контактов, состоит в их описании с помощью расстояний между соответствующими атомами лиганда и рецептора, однако такое приближение и не является физически корректным. Водородные связи описываются эмпирическими геометрическими параметрами (расстояние между донором и акцептором и угол между ними и атомом водорода), а не их энергетическими характеристиками .
Альтернативный подход через статистические оценочные функции заключается в получении основанного на знаниях для взаимодействий из базы данных комплексов белок-лиганд — PDB , и оценки соответствия решения докинга в соответствии с предполагаемым потенциалом .
Существует много программ для теоретического докинга белков. Большая часть работает так: один белок фиксируется в пространстве, а второй поворачивается вокруг него. При этом для каждой конфигурации поворотов производятся оценочные расчеты по оценочной функции. Оценочная функция основана на поверхностной комплементарности, электростатических взаимодействиях, Ван-дер-Ваальсовском отталкивании и так далее. Проблема при этом поиске в том, что вычисления по всему конфигурационному пространству требуют много времени на вычисления, редко приводя к единственному решению .
Несовершенство оценочной функции неизбежно приводит к необходимости оценки предсказательной способности конкретного алгоритма (к примеру AutoDock, ICM) докинга. Для этого необходимы дополнительные экспериментальные данные, к примеру референсная структура. Оценка может быть проведена несколькими способами :
Точность докинга ( англ. Docking accuracy ) — одна из оценок применимости алгоритма, способность алгоритма воспроизводить экспериментальные данные.
Фактор обогащения ( англ. Enrichment factor ) оценивается как способность алгоритма выделить (представить в топе лучших) «истинные» лиганды от «ложных» в выборке, где количество «ложных» много больше количества «истинных». Под «истинными» понимаются лиганды, связывание которых экспериментально доказано, а под «ложными» — лиганды, связывание которых не доказано. Часто проводится анализ ROC -кривой метода .
Способность программ для докинга воспроизводить структуры, полученные методом РСА может быть оценена рядом бенчмаркинг -методов .
В случае малых молекул для сравнительного анализа могут быть взяты специальные референсные наборы, содержащие экспериментальные данные. К примеру, Astex Diverse Set , содержащий структуры белков с лигандами, полученных с помощью рентгеноструктурного анализа или метода Directory of Useful Decoys (DUD) .
В случае докинга пептидов можно использовать Lessons for Efficiency Assessment of Docking and Scoring (LEADS-PEP) .
Последнее время появляется все больше и больше научных статей посвященных виртуальному скринингу и докингу. Однако не стоит слепо доверять их результатам. К наиболее часто возникающим проблемам исследователей можно отнести:
Со стремительным развитием большого количества различных алгоритмов, появляется также проблема выбора наиболее подходящего алгоритма. Наилучшая стратегия выбора заключается в поиске того алгоритма, который был протестирован на подходящей выборке для поставленной задачи и показал оптимальные значения .
В биологии большое количество биохимических процессов протекает на макромолекулярном уровне . Процессы опосредованы белок-белковыми и белок- нуклеиновыми -взаимодействиями. Для изучения такого типа взаимодействий применяют макромолекулярный докинг. Данный метод позволяет предсказать трехмерную структуру изучаемого комплекса в естественной среде. Подобно молекулярному докингу, результатом исследования является набор моделей комплекса (структур), которые далее ранжируются исходя из .
Данный метод позволяет решать большее количество биологических задач .



