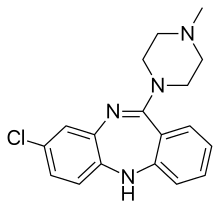
Биохимия (журнал)
- 1 year ago
- 0
- 0



Антагонист ( антагонист рецепторов , рецепторный антагонист ) в биохимии и фармакологии — подтип лигандов к клеточным рецепторам . Лиганд, обладающий свойствами антагониста рецепторов — это такой лиганд, который блокирует, снижает или предотвращает вызываемые связыванием агониста (в том числе эндогенного агониста ) с рецептором физиологические эффекты. Сам же он при этом не обязан (хотя и может ) производить какие-либо физиологические эффекты вследствие своего связывания с рецептором (а по строгому определению, которое подразумевает и включает в себя только нейтральные антагонисты, он даже и не должен производить какие-либо физиологические эффекты сам по себе). Таким образом, антагонисты рецепторов имеют сродство (аффинность) к данному конкретному типу рецепторов, но, исходя из строгого определения, не имеют собственной внутренней агонистической активности по отношению к этому рецептору (вернее она равна нулю), и их связывание лишь нарушает взаимодействие [конкурентных] полных или частичных агонистов с рецептором и предотвращает или ингибирует их функцию и их физиологические эффекты. В равной степени антагонисты рецепторов предотвращают и воздействие на рецептор обратных агонистов . Антагонисты рецепторов опосредуют свои эффекты благодаря связыванию либо с активным сайтом рецептора (так называемым «ортостерическим сайтом» — «правильным местом» связывания), тем же самым, с которым связывается и физиологический эндогенный агонист, либо с аллостерическими сайтами («другими местами связывания» — с которыми могут связываться другие биологически активные по отношению к этому рецептору эндогенные вещества), или же они могут взаимодействовать с рецептором в уникальных участках связывания, которые не являются нормальными участками связывания эндогенных веществ для данного рецептора и не принимают в норме участия в физиологической регуляции активности данного рецептора (часто, однако, обнаружение таких необычных участков связывания предшествует обнаружению в организме их эндогенных лигандов).
Воздействие рецепторного антагониста на рецептор может быть (полностью и быстро) обратимым, труднообратимым или частично и медленно обратимым, или полностью необратимым, в зависимости от продолжительности существования антагонист-рецепторного комплекса. А это, в свою очередь, зависит от природы конкретного антагонист-рецепторного взаимодействия (например, ковалентная связь , как у пиндобинда и феноксибензамина, обычно необратима). Большинство лекарств-антагонистов рецепторов проявляют свои свойства, конкурируя с эндогенными лигандами или субстратами рецептора в структурно строго определённых участках — сайтах связывания — рецепторов.
Клеточные рецепторы — это большие белковые молекулы, которые могут активироваться при связывании с ними эндогенного лиганда (такого, как гормон или нейромедиатор , или цитокин , в зависимости от типа рецепторов), или экзогенного агониста (такого, как лекарство или ). Клеточные рецепторы могут быть трансмембранными , с наружной частью, выступающей с поверхности мембраны клетки , а могут быть внутриклеточными, такими, как ядерные рецепторы (в ядре или на митохондриях или других органеллах клетки). Связывание физиологических эндогенных лигандов (и большинства экзогенных лигандов) с рецептором является результатом нековалентного взаимодействия между лигандом и рецептором, в специфических участках, которые называются «участками связывания» или «сайтами связывания» или «доменами связывания» (также биндинг-сайтами или биндинг-доменами), или «активными сайтами», «активными доменами» данного рецептора. У одного и того же рецептора может иметься несколько активных сайтов (несколько участков связывания) для разных лигандов. Связывание лиганда с рецептором непосредственно регулирует активность рецептора (в частности, связывание агониста с рецептором непосредственно активирует рецептор, вернее повышает вероятность его перехода в активную конфигурацию, облегчает такой переход, делает его более энергетически выгодным, а связывание так называемого «обратного агониста», наоборот, деактивирует или ингибирует рецептор, угнетает его конституциональную встроенную активность, снижает вероятность спонтанной активации рецептора и тем самым стабилизирует его в неактивном состоянии). Активность рецептора также может аллостерически регулироваться с помощью связывания лигандов с другими участками (сайтами) рецептора, называемыми аллостерическими сайтами связывания. Антагонисты опосредуют свои эффекты через взаимодействие с рецепторами благодаря предотвращению воздействия на рецептор как агонистов, так и обратных агонистов, и предотвращению вызывания агонистами и обратными агонистами их соответствующих физиологических эффектов. Это может достигаться через взаимодействие антагониста как с активным сайтом рецептора, так и с одним из его аллостерических сайтов. В дополнение к этому, антагонисты могут взаимодействовать с рецепторами в уникальных участках связывания, в норме не вовлечённых в регуляцию активности рецептора, и проявлять свои эффекты через это взаимодействие.
Термин «антагонист» первоначально применялся в медицине и фармакологии по отношению к совершенно различным профилям фармакологических эффектов лекарств и различным механизмам их антагонистического действия. Тогдашний уровень понимания проблемы и доступные тогда экспериментальные технологии не позволяли провести различение между слабыми или очень слабыми парциальными агонистами, «молчаливыми» (нейтральными) антагонистами и обратными агонистами (это различение бывает довольно затруднительным даже сегодня), и даже о самом существовании таковых подкатегорий антагонистов тогда не подозревали. Более того, во многих случаях тогда не проводилось даже различения между прямым рецепторным антагонизмом (то есть тем, что мы сегодня понимаем под «антагонистом» в контексте фармакологии), и непрямым антагонизмом через воздействие на другие, антагонистически направленные, физиологические процессы или рецепторные структуры, метаболические каскады. То есть термин «антагонист» понимался в физиологическом смысле. В этом смысле, например, «антагонистами» считались адреналин и ацетилхолин (по их клинически противоположному воздействию на сердце и другие органы, а также по их способности угнетать выделение друг друга — о существовании пресинаптических тормозных гетерорегуляторных рецепторов тогда тоже не подозревали). Близкое к современному биохимическое определение термина «рецепторный антагонист», или «антагонист рецепторов» было впервые предложено Аренсом, который также предложил термины «аффинность» и «внутренняя агонистическая активность», в 1954 году и затем усовершенствовано Стивенсоном в 1956 году . Текущее общепринятое определение антагониста рецепторов основывается на рецепторной теории, модели «занятости рецепторов» и на современном (на 2015 год) понимании природы взаимодействий лекарств с рецепторами. Оно сужает изначальное (физиологическое) определение антагонизма до тех соединений, которые проявляют противоположную агонистам («обратную агонистическую») или препятствующую действию агониста («молчаливую антагонистическую») активность по отношению к конкретным белковым молекулам конкретного подтипа рецептора. При этом, согласно определению рецепторного антагонизма, этот антагонизм должен реализовываться непосредственно на уровне самого рецептора как такового. А не, например, на уровне воспрепятствования подозреваемым «антагонистом» работе нисходящего от данного рецептора эффекторного каскада, или на уровне препятствования биосинтезу и экспрессии рецептора, или на уровне препятствования биосинтезу и выделению эндогенного лиганда или ускорения его разрушения, или на уровне воздействия на рецепторы с противоположной направленностью, как в случае адреналина и ацетилхолина.
Первоначально рецепторные системы предполагались вещью двоичной по своей природе. То есть предполагалось, что у рецептора есть (может быть) только два состояния — «активное» и «неактивное», и что промежуточных состояний, промежуточных конфигураций рецептора, не существует (это оказалось не так — промежуточные между «активным» и «неактивным» состоянием конфигурации рецептора существуют ). И предполагалось, что агонисты «включают» рецептор в некое «состояние единицы», то есть что связывание агониста с рецептором вызывает только один , единичный и однозначно определяемый и понимаемый клеточный ответ (это тоже оказалось не так — один и тот же рецептор может в разных ситуациях опосредовать разные внутриклеточные эффекты, и даже одновременно несколько разных и разнонаправленных внутриклеточных эффектов, запуская одновременно несколько разных нисходящих сигнальных каскадов; оказалось также, что одни агонисты более склонны запускать одни нисходящие эффекторные каскады, а другие — другие каскады, так называемое явление функциональной селективности). Более того, в этой упрощённой первоначальной модели предполагалось, что агонисты всегда активируют рецептор, запуская тем самым биохимический механизм изменений внутри клетки (на самом же деле, даже самый высокоэффективный агонист, включая эндогенные агонисты со 100 % эффективностью и недавно открытые суперагонисты, активирует рецептор не всегда , а лишь повышает — и, в случае высокоэффективного агониста, значительно повышает — вероятность перехода рецептора в активированное состояние, делая это состояние, эту конфигурацию более энергетически выгодной). В этой упрощённой модели действие антагониста также предполагалось как простое «выключение», а вернее, предотвращение включения рецептора благодаря связыванию антагониста с ним и предотвращению связывания агониста. Тот факт, что рецепторы могут спонтанно с некоторой вероятностью переходить в активированное состояние даже в отсутствие агониста (то есть обладать некоторой конституциональной активностью) и что антагонист (если он нейтральный) может никак не мешать этой конституциональной активности, или, напротив, может ей мешать и понижать вероятность спонтанной активации рецептора (как в случае обратного агониста) также в этой упрощённой модели не учитывался.
Термин «антагонист» в физиологическом контексте, то есть «физиологический антагонист», «функциональный антагонист» или, иначе, «непрямой антагонист» (вещество, которое производит действие, противоположно направленное действию агониста, но действует на другие рецепторные системы с противоположным эффектом, или даже на ту же систему, но не на рецепторном уровне, а на уровне выше — как в случае блокады биосинтеза лиганда или ускорения его разрушения, или на уровне ниже, как в случае блокады нисходящего от рецептора эффекторного каскада) — также всё ещё употребителен. Примером такого «физиологического», или «непрямого» функционального антагонизма может служить тот факт, что гистамин и ацетилхолин снижают артериальное давление , вызывая вазодилатацию через, соответственно, гистаминовые и ацетилхолиновые рецепторы, в то время как адреналин повышает артериальное давление, вызывая вазоконстрикцию через адренорецепторы. Другой пример «непрямого» антагонизма — тот факт, что тригексифенидил , холиноблокатор, снижает экстрапирамидные побочные эффекты галоперидола , D2-блокатора .
Наше понимание механизмов вызываемой лекарствами и эндогенными веществами активации рецепторов и рецепторной теории и современное биохимическое определение рецепторного антагониста продолжает эволюционировать и совершенствоваться и сегодня. Примитивное понимание состояния активации рецептора как двузначной логики («ноль» — «выключено» или «единица» — «активировано») уступило место современной многозначной логической модели, признающей существование множества промежуточных пространственных конфигураций рецептора. Примитивное представление о 100-процентной вероятности активации рецептора после связывания с агонистом и нулевой вероятности спонтанной активации рецептора (в отсутствие агониста) уступило место современной вероятностной модели, согласно которой рецепторный белок постоянно спонтанно осциллирует между множеством «неактивных» и «активных» конфигураций, с некоторой вероятностью пребывая в каждый момент времени в том или ином состоянии, то есть имеет некий ненулевой базальный, встроенный уровень конституциональной активности (зависящий от вероятности спонтанной активации конкретного белка конкретного подтипа рецепторов в конкретном микроокружении), а агонист не со 100 % вероятностью «включает» белок, а лишь повышает вероятность такого «включения», делает его более энергетически выгодным. Открытие явления конституциональной внутренней активности рецепторов привело к открытию явления обратного агонизма и к переопределению многих «антагонистов рецепторов» как обратных агонистов (классический пример — антигистаминные средства ). Открытие того факта, что эндогенные лиганды не со 100 % вероятностью активируют рецептор, привело к открытию так называемых «суперагонистов» — агонистов, более эффективно активирующих рецептор, чем эндогенные лиганды (что ранее считалось невозможным). Открытие явления функциональной селективности и того, что наиболее энергетически выгодные и, соответственно, наиболее вероятные конфигурации рецептора зависят от конкретного лиганда (являются лиганд-специфичными) и что разные конфигурации рецептора могут по-разному (дифференцированно) активировать разные связанные с данным рецептором нижележащие сигнальные каскады и системы вторичных посредников привело к пониманию возможности создания лекарств, которые будут селективно (избирательно) активировать некоторые из нижележащих сигнальных каскадов рецептора и не активировать другие, и, соответственно, будут лишены побочных эффектов «классических» агонистов и антагонистов. Это также означает, что рецепторная эффективность (внутренняя агонистическая активность) того или иного агониста или антагониста может зависеть от микроокружения данного рецептора, от того, где именно, в какой именно ткани и в каких именно клетках данной ткани экспрессирован данный рецептор. Это меняет наши первоначальные представления о том, что тот или иной уровень рецепторной эффективности (внутренней агонистической активности) есть характеристическое свойство самого лекарства, не зависящее от свойств конкретного рецептора в конкретном организме и даже в конкретной ткани и клетке. И открывает путь к синтезу лекарств, которые селективно блокируют или стимулируют рецепторы в одних тканях, в то же время сравнительно мало влияя на аналогичные рецепторы в других тканях. Хорошим примером являются атипичные антипсихотики , которые сравнительно сильно блокируют дофаминовые рецепторы в мезолимбической и мезокортикальной областях головного мозга , где такая блокада нужна и обеспечивает полезный антипсихотический эффект, и сравнительно мало блокируют те же дофаминовые рецепторы в нигростриарной системе, гипоталамусе и гипофизе , где такая блокада вредна и вызывает экстрапирамидные побочные явления и повышение пролактина . Другим характерным примером тканеспецифической активации/блокады рецепторов в зависимости от конкретной ткани является селективный модулятор эстрогеновых рецепторов ралоксифен, стимулирующий эстрогеновые рецепторы в костях (предотвращает остеопороз ) и в гипоталамусе (снимает явления климакса ) и одновременно блокирующий те же самые рецепторы в молочных железах , снижая вероятность рака молочной железы как по сравнению с классической эстроген-заместительной терапией (полными агонистами эстрогеновых рецепторов), так и по сравнению с отсутствием лечения.
Согласно строгому определению термина, «истинные» антагонисты рецепторов проявляют нулевую внутреннюю агонистическую активность (то есть обладают нулевой рецепторной эффективностью, или, иначе говоря, не обладают способностью активировать рецепторы, с которыми связываются, даже в самой малой степени). Однако, связываясь с рецепторами, «истинные» антагонисты рецепторов предотвращают воздействие на эти же рецепторы агонистов, обратных агонистов и частичных агонистов. В функциональных исследованиях антагонистов кривая «доза-эффект» позволяет измерить и графически представить способность антагониста рецепторов в определённом диапазоне клинически применяемых концентраций (как правило, наномолярных) ингибировать или предотвращать действие агониста. В действительности существует очень мало «истинных» антагонистов рецепторов, внутренняя агонистическая активность которых по отношению к данному подтипу рецепторов строго равна нулю — как правило, все они на самом деле являются либо слабыми или очень слабыми частичными агонистами (внутренняя агонистическая активность менее 10-20 %, или вообще очень мала и не обнаруживается существующими методами исследования, но это вовсе не обязательно означает, что она равна нулю), либо обратными агонистами.
Молярная активность антагониста рецепторов обычно определяется как его половинная эффективная концентрация, или так называемая величина EC 50 . Величина EC 50 для данного антагониста рецепторов вычисляется путём определения концентрации антагониста рецепторов, вызывающей 50-процентное ингибирование максимального биологического ответа на соответствующий агонист тех же рецепторов. Определение EC 50 полезно для сравнения молярной активности разных антагонистов данного рецептора с равной или близкой (сходной) внутренней агонистической активностью. Однако для того, чтобы такое прямое сравнение двух антагонистов рецепторов по их молярной активности было возможным и было корректным, необходимо, чтобы и форма кривой «доза-эффект» у обоих лекарств была близкой или сходной, а это не всегда так. В противном же случае такое сравнение будет либо невозможным, либо некорректным. Чем меньше значение EC 50 , то есть чем меньше доза, требуемая для 50 % ингибирования максимального биологического ответа на агонист в исследованиях, тем выше молярная активность данного антагониста рецепторов, и тем ниже доза и концентрация в крови лекарства, требуемая для достижения ингибирования биологического ответа на агонист и в живом организме.
Более высокая молярная активность антагониста, то есть возможность применять более низкие дозы и концентрации антагониста рецепторов для достижения того же самого эффекта, что и у менее активного антагониста, как правило, сопряжена с большей избирательностью антагониста по отношению именно к данному подтипу рецепторов, меньшей метаболической нагрузкой на печень и меньшей выделительной нагрузкой на органы выделения ( почки , кишечник и др.), меньшей токсичностью и меньшими побочными эффектами. И напротив, менее активные антагонисты рецепторов часто являются «грязными» ( dirty drug ) в смысле недостаточной избирательности по отношению к нужному подтипу рецепторов и количества вызываемых ими побочных эффектов и токсичности. Хорошим примером здесь является сравнение малоактивных типичных антипсихотиков (например, хлорпромазина , терапевтические дозы которого при психозах измеряются сотнями миллиграммов) с более высокоактивными соединениями, такими, как галоперидол или перфеназин (терапевтические дозы которых при тех же состояниях измеряются десятками миллиграммов). Хлорпромазин обладает значительной гепатотоксичностью. А кроме того, он помимо связывания с D 2 дофаминовыми рецепторами, опосредующими его клинически полезный антипсихотический эффект, также связывается со множеством рецепторов и белков, связывание с которыми нежелательно и вредно и ничего, кроме побочных эффектов, не приносит. Например, способность хлорпромазина связываться с α 1 -адренорецепторами опосредует такие его нежелательные побочные эффекты, как гипотензия , тахикардия , ортостатические обмороки и коллапсы. Способность хлорпромазина связываться с гистаминовыми H 1 -рецепторами опосредует нежелательную сонливость и седацию, повышение аппетита и прибавку массы тела. Способность связываться с M-холинорецепторами опосредует сухость во рту, задержку мочеиспускания, запоры. Способность хлорпромазина связываться с моноаминовыми транспортерами и вызывать истощение депо моноаминов — опосредует нередко вызываемую им депрессию. Эти свойства у более активных по отношению к D 2 -рецепторам типичных антипсихотиков, таких, как галоперидол, перфеназин, выражены гораздо слабее, при большей антипсихотической активности, вследствие как раз их более высокой избирательности по отношению к «нужным» D 2 -рецепторам и меньшего связывания с другими, «ненужными» типами рецепторов, а также меньшей метаболической нагрузки на печень в клинически эффективных дозах. Другим характерным примером может служить сравнение трициклических антидепрессантов, таких, как амитриптилин , имипрамин (эффективные дозы которых также измеряются сотнями миллиграммов) с СИОЗС (эффективные дозы которых измеряются десятками миллиграммов) — у последних также намного выше избирательность и намного меньше побочных эффектов.
Высокая молярная активность и высокая избирательность антагониста по отношению к нужному типу рецепторов важны также и при использовании его в исследовательских целях, например в качестве радиоактивного лиганда исследуемых рецепторов при ПЭТ . Возможность использовать меньшее количество радиоактивного лиганда и получить тот же процент занятости рецепторов в силу более высокой потентности лиганда означает меньшую лучевую нагрузку от ПЭТ. А более высокая селективность — означает более корректные результаты исследования (не будут ложно помечены и «засвечены» радиолигандом другие, отличные от подвергающегося исследованию, типы рецепторов).
Таким образом, синтез более активных и более избирательных, а значит и менее токсичных, по сравнению с существующими, антагонистов различных типов рецепторов является актуальной задачей современной экспериментальной и клинической фармакологии.
Степень сродства (аффинность) антагониста по отношению к его сайту связывания (K i ), то есть его способность связываться со специфическим участком рецептора, предопределяет продолжительность ингибирования им эффектов агонистов. Степень сродства антагониста по отношению к данному сайту связывания данного подтипа рецепторов может быть определена экспериментально с использованием метода регрессии Шильда, или, в случае конкурентных антагонистов, с помощью изучения связывания меченого радиоактивной меткой лиганда с использованием уравнения Ченджа-Прусоффа. Метод регрессии Шильда может быть использован для определения природы антагонизма как конкурентного или неконкурентного. Определение K i антагониста этим методом также не зависит от сродства к рецептору, от величины внутренней агонистической активности или от молярной концентрации использованного агониста. Однако для использования этого метода необходимо, чтобы в исследуемой системе было предварительно достигнуто динамическое равновесие (эквилибриум). Кроме того, следует принимать во внимание влияние эффекта десенситизации рецепторов под воздействием агониста, и, напротив, сенситизации их под влиянием антагониста, на достижение эквилибриума. Кроме того, методом регрессии Шильда не может быть проанализирована и достоверно установлена степень сродства к рецепторам веществ, проявляющих в исследуемой системе два или более различных эффекта, как, например, конкурентные антидеполяризующие миорелаксанты, которые не только конкурентно ингибируют связывание агониста (ацетилхолина) с мембраной нервно-мышечного синапса, но и непосредственно блокируют ионные каналы. Особые трудности возникают в том случае, если эти физиологически различные на субклеточном уровне эффекты не отличимы или трудно отличимы друг от друга функционально с использованием выбранного метода измерения агонистического эффекта (как определить — почему мышечная клетка расслабилась — по причине ли блокады ацетилхолинового рецептора, или по причине блокады ионного канала?). Метод регрессии Шильда заключается в сравнении изменений, вызываемых добавлением данной дозы конкурентного антагониста, в эффективной концентрации агониста (EC 50 ) по сравнению с EC 50 агониста в отсутствие антагониста и масштабировании получаемых величин EC 50 по отношению к исходной EC 50 в отсутствие антагониста (вычислении относительной дозы — dose ratio ). Изменяя дозу антагониста, можно изменить EC 50 агониста. Таким образом, в методе регрессии Шильда рисуется график, по одной оси которого — логарифм относительной дозы агониста, а по другой — логарифм концентрации антагониста для достаточно широкого диапазона его концентраций. Сила сродства антагониста к рецептору (аффинности), или значение K i , в этом случае находится там, где аппроксимированная линия графика регрессии Шильда пересекает ось X.
В то время как в методе регрессии Шильда в экспериментах для определения величины K i варьируется концентрация антагониста , для определения величины K i по методу уравнения Ченджа-Прусоффа используется другой метод — варьируется концентрация агониста . Сродство к рецепторам для конкурентных агонистов и антагонистов в этом случае определяется уравнением Ченджа-Прусоффа из сдвига в эффективной ингибирующей концентрации антагониста (IC 50 ), который происходит при варьировании концентрации агониста в процессе конкурентного антагонизма. Уравнение Ченджа-Прусоффа позволяет учесть влияние изменения концентраций агониста и сродства агониста к рецептору на десенситизацию рецепторов и на угнетение их активности, производимое антагонистами. Поскольку в физиологических условиях в живом организме — как в норме, так и при патологии — обычно изменяется концентрация именно физиологического агониста , причём в довольно широких пределах, в то время как изменять концентрацию антагониста в крови мы можем лишь в достаточно узких пределах от нуля до некоторого разумного предела (чрезмерное повышение дозы чревато потерей избирательности антагониста по отношению к нужному типу рецепторов и различными побочными эффектами), то уравнение Ченджа-Прусоффа, теоретически, позволяет получить более близкие к реальному сродству вещества к рецепторам в реальном живом организме значения K i , чем уравнение регрессии Шильда.
Конкурентные антагонисты обратимо связываются с рецепторами в том же самом активном сайте связывания, что и физиологический эндогенный лиганд-агонист этого рецептора, но не вызывают активации рецептора (или вызывают его с пренебрежимо малой вероятностью, намного меньшей, чем эндогенный агонист, как это имеет место в случае «слабых частичных агонистов», которые также могут являться конкурентными антагонистами в физиологических условиях). Физиологические (и иные) агонисты и конкурентные антагонисты в этом случае «конкурируют» за связывание с одним и тем же активным сайтом рецепторов. После того, как конкурентный антагонист свяжется с активным сайтом рецептора, он предотвращает связывание с ним же агониста (на то время, пока он сам остаётся связанным с ним, то есть не диссоциировал из связи с ним). Однако конкурентный антагонист не может ни «вытеснить» уже связавшийся с рецептором агонист из связи (до тех пор, пока сам агонист не диссоциирует из этой связи — а вероятность этого события определяется его кинетикой, в частности константой диссоциации физиологического агониста), ни предотвратить воздействие уже связавшегося агониста на клетку (активацию рецептора). Конечный результат конкуренции агонистов и антагонистов — и таким образом конечный уровень активности рецепторной системы — определяется соотношением молярных концентраций, относительных аффинностей и относительной внутренней агонистической активности агонистов и антагонистов. Поскольку высокие концентрации конкурентного антагониста повышают процент занятости рецепторов этим антагонистом, для достижения того же самого процента занятости рецепторов агонистом в этих условиях — и получения того же самого физиологического ответа — потребуются более высокие концентрации агониста, и наоборот — при более высоких концентрациях агониста требуется больше конкурентного антагониста для функциональной «блокады» рецепторов. В функциональных исследованиях конкурентные антагонисты вызывают параллельный сдвиг кривой «доза агониста-эффект» вправо, без изменения максимальной величины физиологического ответа (в отличие от неконкурентных , а также необратимых антагонистов, которые изменяют именно максимальную величину физиологического ответа).
Антагонист рецепторов интерлейкина-1 является примером конкурентного антагонизма. Эффект конкурентного антагониста может быть преодолён увеличением концентрации агониста. Часто (хотя и не всегда) конкурентные антагонисты обладают химической структурой, очень сходной с химической структурой агонистов тех же самых рецепторов (физиологического агониста или других уже известных агонистов). Если же не наблюдается сходства химической структуры, то, во всяком случае, обычно наблюдается сходство пространственной структуры именно того участка молекулы антагониста, который непосредственно связывается с активным сайтом рецептора (более того, как можно предположить, это сходство пространственной структуры необходимо для взаимодействия с активным сайтом рецептора).
Термин «неконкурентный антагонизм» используется для описания двух разных феноменов: в одном случае неконкурентный антагонист связывается с ортостерическим активным сайтом рецептора (с тем же самым, с которым связывается физиологический агонист), а в другом случае он связывается с аллостерическим сайтом рецептора (то есть другим, не тем же самым, с которым связывается физиологический агонист). И хотя механизм антагонистического действия различен в обоих случаях, они оба называются «неконкурентным антагонизмом», поскольку конечный результат воздействия антагониста в обоих случаях функционально очень похож. В отличие от конкурентных антагонистов, которые конкурируют с агонистами за занятость рецепторов и сдвигают кривую доза-эффект вправо, влияя на количество агониста, необходимое для получения максимального физиологического ответа (чем больше доза или концентрация конкурентного антагониста, тем больше нужно агониста, чтобы вызвать тот же самый физиологический ответ), но никак не влияют на саму величину максимального физиологического ответа («вершину кривой доза-эффект»), неконкурентные антагонисты уменьшают величину максимального физиологического ответа, который может быть получен при любом сколь угодно большом количестве агониста. Это свойство и даёт им название «неконкурентные антагонисты», поскольку их эффект не может быть «уничтожен», обнулен или скомпенсирован увеличением количества агониста, сколь бы велико это увеличение ни было. В биологических системах, предназначенных для изучения влияния тех или иных антагонистов на рецепторы, неконкурентные антагонисты вызывают уменьшение «плато» (максимального значения кривой «доза агониста-эффект»), и, в некоторых случаях, также сдвиг кривой вправо. Сдвиг кривой вправо происходит вследствие наличия во многих биологических рецепторных системах так называемого «рецепторного резерва» («запасных рецепторов») , и ингибирование агонистического ответа под влиянием неконкурентного антагониста происходит только тогда, когда истощится (израсходуется) этот рецепторный резерв.
Антагонист, который связывается с активным сайтом рецептора, обычно оказывается и называется «неконкурентным», если связь между активным сайтом рецептора и антагонистом не ковалентная, но по тем или иным причинам очень прочная и трудно разрывается или вообще не разрывается в течение длительного времени (превышающего время эксперимента), что создаёт у исследователя или практического врача иллюзию необратимой инактивации рецептора. Однако такое использование термина не идеально и часто приводит к путанице, поскольку термин «труднообратимый конкурентный антагонизм» более удачен, лучше описывает сущность явления и не создаёт путаницы как с понятием «необратимый антагонизм» (подразумевающим ковалентную связь антагониста с рецептором и его необратимую порчу, требующую биосинтеза новых рецепторов взамен деградировавших), так и с вторым значением понятия «неконкурентный антагонизм», подразумевающим связывание антагониста с аллостерическим сайтом рецептора и обычно обратимую (хотя также иногда трудно обратимую или вовсе необратимую) аллостерическую модификацию его конфигурации таким образом, что в этой конфигурации предотвращается связывание агониста.
Второе значение термина «неконкурентный антагонист» относится к антагонистам, связывающимся с аллостерическим сайтом рецептора (то есть не с тем же самым сайтом, с которым связывается физиологический агонист). Эти антагонисты связываются с рецептором в ином месте, чем физиологический агонист, и оказывают своё действие на рецептор через этот сайт (называемый аллостерическим сайтом связывания рецептора ). Поэтому они не конкурируют с агонистами за связывание с активным (ортостерическим) сайтом рецептора, и их эффективность не зависит поэтому от концентрации агониста в среде. Связавшийся с аллостерическим сайтом рецептора антагонист производит процесс, называемый «аллостерической модификацией рецептора» — а именно, в нашем случае (антагонизм) — предотвращает или снижает вероятность конформационных изменений рецептора, требуемых для его активации, при связывании агониста (то есть агонист — может свободно связываться с рецептором, но активации рецептора при этом не произойдёт или она существенно менее вероятна), или же меняет конфигурацию рецептора таким образом, что затрудняется или становится вообще невозможным связывание агониста (меняется конфигурация активного сайта рецептора). Так, например, циклотиазид оказался обратимым неконкурентным аллостерическим антагонистом метаботропного глутаматного рецептора подтипа 1 (mGluR 1 ).
Термин «бесконкурентные антагонисты» ( uncompetitive antagonist ) отличается по значению от термина «неконкурентные антагонисты» ( non-competitive antagonist ). Этот термин применяется по отношению к антагонистам, которые сами по себе не связываются с неактивной формой рецептора (то есть в отсутствие связывания агониста с рецептором), однако способны связываться с активной (активированной предварительным связыванием агониста) формой рецептора в специфическом аллостерическом сайте связывания (другом, не том, с которым связывается агонист), эффективно предотвращая тем самым активацию рецептора агонистом и его переход в активную конфигурацию (или, вернее, снижая вероятность такого перехода под влиянием уже связавшегося агониста). То есть такие бесконкурентные антагонисты для своего связывания с рецептором требуют предварительной активации рецептора агонистом. Бесконкурентный тип антагонизма даёт характерный «парадоксальный» (противоречащий привычной логике рецептор-лигандных взаимодействий) кинетический профиль, в котором явление выглядит так, что «одно и то же количество бесконкурентного антагониста эффективнее блокирует рецепторную активацию при более высокой концентрации агониста, чем при более низких концентрациях». Одним из примеров такого бесконкурентного антагонизма является мемантин , лекарство, применяемое в лечении болезни Альцгеймера — он является бесконкурентным антагонистом NMDA-рецепторов. Важным преимуществом такого подхода является то, что этот механизм обеспечивает не простую «блокаду» тех или иных физиологических функций, обеспечиваемых агонистом, а тонкую регуляцию — при меньшей концентрации физиологического агониста происходит меньшая блокада бесконкурентным антагонистом (так как меньше активированных рецепторов), при более высокой концентрации физиологического агониста то же количество, та же доза бесконкурентного антагониста обеспечивает более высокую степень блокады, эффективно ограничивая её сверху, но не мешая некоему базовому низкому уровню активации. Это обеспечивает меньшее количество и другой спектр побочных эффектов мемантина по сравнению с такими «традиционными» NMDA-антагонистами, как кетамин , и другую сферу применения мемантина.
«Молчаливые», или нейтральные, антагонисты — это такие конкурентные антагонисты данного типа рецепторов, которые имеют строго нулевую внутреннюю агонистическую активность, то есть нулевую способность активировать рецептор (в отличие от слабых частичных агонистов, у которых такая способность мала, но всё же не строго равна нулю), но также и не препятствуют конституциональной внутренней активности рецептора, не снижают её (то есть не снижают частоту «спонтанной активации» рецептора) и не обладают собственными, отличными от блокирования связывания рецептора с агонистом, физиологическими эффектами по отношению к данной рецепторной системе. В некотором смысле именно «молчаливые антагонисты» являются «истинными», «настоящими» антагонистами, в первоначальном значении слова (которое употреблялось до открытия конституциональной активности рецепторов и факта существования обратных агонистов, а также до установления того факта, что многие из лекарств, считавшихся «антагонистами» того или иного типа рецепторов, на самом деле являются либо слабыми частичными агонистами, либо обратными агонистами).
Этот термин был создан именно для того, чтобы отличать «истинные» (полностью неактивные) по отношению к данному конкретному типу рецепторов, антагонисты — от слабых парциальных агонистов и от обратных агонистов.
Однако на практике «истинных» нейтральных или молчаливых антагонистов очень мало — очень редко внутренняя агонистическая активность того или иного соединения действительно строго равна нулю. Подавляющее большинство соединений, считающихся «нейтральными антагонистами» являются либо слабыми и очень слабыми частичными агонистами (с внутренней агонистической активностью менее 10-20 %), либо (слабыми) обратными агонистами. Во многих экспериментальных биологических системах невозможно или очень затруднительно отличить слабые частичные агонисты и «истинные» нейтральные антагонисты, а также различить нейтральные антагонисты и обратные агонисты (особенно в случае слабого обратного агонизма). И даже в случаях, когда кажущаяся внутренняя агонистическая активность некоего соединения в некоем предположительно «высокоточном» эксперименте, уточняющем наши первоначальные представления, оказалась действительно равной нулю — это, на самом деле, значит лишь, что она меньше порога чувствительности данного экспериментального метода (например, условно говоря, +0.1 % или −0.1 %).
Частичные агонисты тех или иных рецепторов (также называемые парциальными агонистами) — вещества, которые могут отличаться от эндогенного агониста (максимальная амплитуда вызываемого которым физиологического ответа клеток принимается за 100 % согласно определению) по максимальной амплитуде вызываемого ими физиологического эффекта в меньшую сторону, при максимально возможной занятости данного типа рецепторов данным [частичным] агонистом. Хотя частичные агонисты, как следует из их названия, являются разновидностью агонистов по отношению к данному типу рецепторов, они могут действовать как конкурентные антагонисты по отношению к этому же типу рецепторов в присутствии полного агониста (в частности, в присутствии физиологического, эндогенного агониста) или в присутствии более сильного и более эффективного (имеющего более высокую внутреннюю агонистическую активность ) частичного агониста. Происходит это потому, что частичные агонисты конкурируют с полным агонистом, в частности с физиологическим эндогенным агонистом (или с более сильным частичным агонистом) за занятость рецепторов. Тем самым частичный агонист, в присутствии полного агониста (в частности, в присутствии физиологического агониста) или в присутствии более сильного частичного агониста, приводит к меньшей степени активации рецепторов и меньшему максимальному физиологическому ответу, в сравнении с воздействием только полного агониста (например, физиологического агониста) или только сильного частичного агониста. На практике, многие лекарства, обычно считающиеся «антагонистами» тех или иных рецепторов, являются слабыми частичными агонистами (с внутренней агонистической активностью, не превышающей 10-20 % от активности эндогенного агониста). Часто слабый частичный агонист (с активностью меньшей 10-20 %) вообще бывает невозможно отличить от истинного «молчаливого» или нейтрального антагониста в условиях эксперимента, в то время как сильный частичный агонист (с активностью, превышающей 70-90 %) точно так же бывает невозможно отличить от «истинного» полного агониста. Клиническая эффективность и польза от существования частичных агонистов заключается в их способности повышать активность «недостимулированных» (дефицитарных, страдающих от недостаточной агонистической стимуляции) систем, и в то же время эффективно блокировать и предотвращать чрезмерную, избыточную и вредную агонистическую стимуляцию, проистекающую от повышенного уровня эндогенных агонистов.
Так, например, арипипразол при шизофрении одновременно повышает активность дофаминергических систем в префронтальной коре, где она у больных шизофренией понижена, и тем самым эффективно уменьшает негативную симптоматику и когнитивные нарушения у больных шизофренией, и в то же время снижает чрезмерно повышенную активность дофаминергических систем в мезолимбической и мезокортикальной областях, где она у больных шизофренией повышена, и тем самым эффективно устраняет бред и галлюцинации. Экспозиция рецепторов с высоким уровнем частичного агониста (такого, как арипипразол по отношению к дофаминовым D2 рецепторам) гарантирует, что данная рецепторная система будет иметь постоянный, но достаточно невысокий, уровень активности, вне зависимости от того, высокая или низкая концентрация эндогенного физиологического агониста присутствует в данном конкретном участке мозга. Кроме того, частичные агонисты, вызывая меньшую степень функциональной блокады (снижения активности) рецепторной системы, чем «молчаливые» антагонисты, и меньшую степень стимуляции рецепторов, чем полные агонисты, обычно имеют меньше побочных эффектов (так как происходит менее грубое вмешательство в работу данной конкретной рецепторной системы). Примером может служить тот же арипипразол, вызывающий значительно меньшее количество экстрапирамидных побочных эффектов и меньшее повышение уровня пролактина , чем типичные антипсихотики наподобие трифлуоперазина и галоперидола , являющиеся «молчаливыми антагонистами» D2-рецепторов.
Другим типичным примером успешного использования принципа частичного агонизма с целью уменьшения побочных эффектов является создание β-адреноблокаторов , обладающих частичной внутренней симпатомиметической активностью и, вследствие этого, вызывающих меньший бронхоспазм , меньшую брадикардию (особенно мало влияя на частоту сердечных сокращений в покое, но эффективно ограничивающих тахикардию при физической нагрузке, стрессе , тревоге ), меньшую периферическую вазоконстрикцию (а иногда даже проявляющих вазодилатирующие свойства), реже вызывающих депрессию и меньше влияющих на обмен веществ , по сравнению с β-адреноблокаторами без внутренней симпатомиметической активности («молчаливыми» β-адреноблокаторами). Кроме того, также принято считать, что более «сбалансированные» частичные агонисты менее склонны вызывать развитие или запуск адаптивных, «ускользающих» от внешнего агонистического или антагонистического воздействия, контр-регуляторных механизмов поддержания гомеостаза , таких, как десенситизация (даунрегуляция) рецепторов при воздействии полного агониста или сильного частичного агониста или, наоборот, сенситизация (апрегуляция) рецепторов при воздействии очень слабого частичного агониста, «молчаливого» антагониста или обратного агониста.
Таким образом, на больших промежутках времени более «сбалансированные» частичные агонисты могут оказываться эффективнее, хотя на коротком промежутке времени, до включения контррегуляторных механизмов, полный агонист, сильный частичный агонист или «молчаливый антагонист/обратный агонист» может оказаться — и часто оказывается — эффективнее «сбалансированного» частичного агониста в вызывании соответствующих (агонистических или антагонистических) эффектов максимальной силы. Примером является бупренорфин , сбалансированный частичный агонист μ-опиоидных рецепторов , который проявляет сравнительно слабую морфиноподобную активность, однако при этом он не только вызывает меньшее угнетение дыхания, оказывает меньшее воздействие на сердечно-сосудистую систему (меньшую степень брадикардии и гипотензии ), меньше запоров по сравнению с морфином (это в целом типично для частичных агонистов — давать меньше побочных эффектов, как описывалось выше), но и вызывает меньшую наркотическую зависимость , менее выраженный абстинентный синдром и меньшую степень гипералгезии . Это связывают именно со способностью бупренорфина, как относительно слабого, «сбалансированного» частичного агониста, вызывать меньшую степень десенситизации опиоидных рецепторов при хроническом применении. Это даже позволяет применять бупренорфин как для детоксикации опиоидных наркоманов и купирования опиоидной абстиненции, так и для длительной поддерживающей опиоидной заместительной терапии неизлечимых наркоманов, как альтернативу метадону . Более того, это позволяет применять бупренорфин вне сфер наркологии и купирования болевого синдрома, как средство лечения резистентных форм депрессий — применение, которое было бы невозможным, если бы бупренорфин обладал сильными опиоидными агонистическими свойствами и, подобно другим сильным опиоидным агонистам, сильно десенситизировал опиоидные рецепторы, сильно угнетал биосинтез эндорфинов и вызывал сильную зависимость и выраженный абстинентный синдром.
Так называемые « обратные агонисты » (или, иначе, «инверсные агонисты» — inverse agonist ) могут вызывать в целостном живом организме эффекты, внешне довольно сходные с эффектами «молчаливых» или «нейтральных» антагонистов (просто молча блокирующих, предотвращающих действие в организме всегда присутствующих в нём в норме физиологических концентраций агониста). Однако на клеточном уровне они вызывают совершенно определённый — принципиально отличный, хотя и не во всякой экспериментальной технике легко отличимый — от эффекта «молчаливых антагонистов» — каскад нисходящих эффекторных реакций, запускаемых их связыванием с рецептором и соответствующим изменением конфигурации рецептора (его инактивацией, снижением его базальной, конституциональной активности, то есть снижением вероятности его спонтанной активации, спонтанного перехода в активированное состояние). Этот каскад нисходящих эффекторных реакций в целом ведёт к проявлению на уровне отдельных клеток физиологических эффектов, в целом противоположных обычно наблюдаемым при воздействии на клетки агонистов (хотя могут наблюдаться также и дополнительные эффекты, не сводимые к простой противоположности эффекту агонистов).
Таким образом, для любых типов рецепторов, которые «от природы» по своему устройству имеют некий определённый базальный уровень конституциональной «внутренней активности» (не зависящей от присутствия или отсутствия агониста), потенциально могут существовать (и нередко уже открыты и описаны) обратные агонисты, которые не только «молча» блокируют, предотвращают связывание агонистов с рецепторами и воздействие агониста, но и ингибируют, угнетают базальную конституциональную активность рецептора. Многие лекарства, ранее традиционно классифицировавшиеся просто как «антагонисты» (и считавшиеся «нейтральными» или «молчаливыми» антагонистами), в настоящее время либо уже переклассифицированы, либо находятся в процессе переклассификации в обратные агонисты, в связи с открытием явления конституциональной внутренней активности рецепторов (которое было ранее неизвестно) и их способности угнетать её. Так, в частности, антигистаминные средства, первоначально классифицировавшиеся как антагонисты гистаминовых рецепторов подтипа H 1 , в настоящее время переклассифицированы как обратные агонисты этого же рецептора.
Возможность наличия у обратных агонистов дополнительных физиологических эффектов, не сводимых к простой противоположности эффектам агонистов (запуск внутриклеточных сигнальных каскадов, отличных от тех, что запускаются «в норме» конституционально активным рецептором, как при связывании агониста, так и в его отсутствие, или при связывании нейтрального антагониста) делает задачу прицельной разработки обратных агонистов различных типов рецепторов одной из интересных задач современной фармакологии. В то же время угнетение базальной конституциональной активности рецептора, очевидно, логически может приводить к более серьёзным побочным эффектам как на уровне клетки, так и на уровне организма в целом, чем простое «выключение» или блокирование рецептора (простое предотвращение его связывания с агонистом).
Большинство антагонистов рецепторов являются обратимыми антагонистами, которые, подобно большинству агонистов, связываются с рецептором и отсоединяются от рецептора с определёнными вероятностями и через определённые промежутки времени, определяемые кинетикой связывания рецептора с лигандом.
Однако существуют так называемые необратимые антагонисты. Необратимые антагонисты ковалентно связываются с рецептором-мишенью, необратимо меняя его пространственную конфигурацию и тем самым необратимо инактивируют его. Необратимые антагонисты, как правило, не могут быть ферментативно удалены из связи с рецептором. Таким образом, продолжительность физиологического воздействия необратимого антагониста определяется не традиционной кинетикой связывания рецептора с лигандом, а скоростью оборота рецепторов — скоростью процесса физиологического «выведения из строя» и удаления с поверхности мембраны клетки старых, «деградировавших» от времени рецепторов и скоростью биосинтеза клеткой и выведения на поверхность клеточной мембраны новых рецепторов взамен старых, деградировавших. Примером необратимого антагониста α-адренорецепторов является феноксибензамин, который ковалентно и необратимо связывается с α-адренорецепторами, предотвращая тем самым связывание с ними адреналина и норадреналина . Инактивация рецепторов необратимым агонистом обычно приводит к снижению или уменьшению максимально возможного физиологического ответа на максимальную агонистическую стимуляцию («уплощение» кривой доза-эффект в зависимости от концентрации агониста, уменьшение её максимума). Помимо этого, в системах, в которых есть рецепторный резерв, также может наблюдаться сдвиг кривой «доза-эффект» вправо, подобный сдвигу кривой вправо, наблюдаемому при воздействии конкурентных антагонистов. Отмывание культуры клеток, которые подверглись воздействию антагониста, от остатков антагониста, обычно позволяет различить воздействие неконкурентного (но обратимого) антагониста от воздействия необратимого антагониста, поскольку действие неконкурентных антагонистов кратковременно и обратимо, и после отмывания клеток от антагониста эффективность воздействия на них агониста восстанавливается, чего не происходит при воздействии необратимых антагонистов.
Действие необратимых конкурентных антагонистов также основано на конкуренции антагониста с агонистом за рецептор. Однако скорость формирования ковалентных связей между рецепторами и таким антагонистом зависит от аффинности и химической реактивности конкретного необратимого антагониста. Для некоторых необратимых конкурентных антагонистов, может наблюдаться конкретный ограниченный во времени период, когда они ведут себя как обычные (обратимые) конкурентные антагонисты (могущие иметь или не иметь ту или иную базальную внутреннюю агонистическую активность), и свободно связываются с рецептором и так же свободно диссоциируют из связи с рецептором, со скоростями и вероятностями, определяемыми традиционной кинетикой связывания рецептора с лигандом. Однако с момента, когда сформировалась необратимая ковалентная связь, рецептор подвергается необратимой деактивации и функциональной деградации. Также как и для неконкурентных обратимых антагонистов и для неконкурентных необратимых антагонистов, в эксперименте для них может наблюдаться сдвиг кривой «доза-эффект» вправо. Однако в целом обычно наблюдается как уменьшение скорости нарастания кривой (первой производной), так и уменьшение максимума кривой.