Большой бинокулярный телескоп
- 1 year ago
- 0
- 0

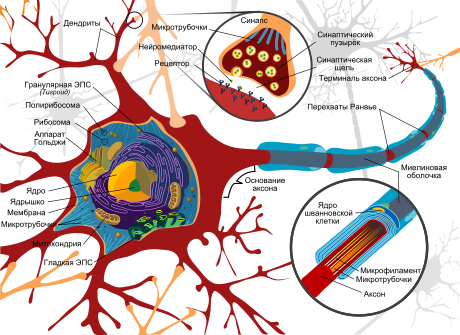
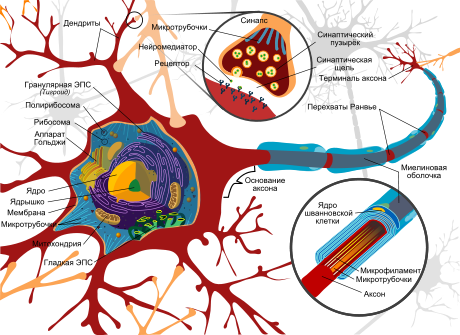
Бинокулярные нейроны — это нейроны зрительной системы , отвечающие на стимуляцию обеих сетчаток глаз .
У бинокулярных нейронов обнаружены большие вариации в относительной силе ответов на стимуляцию сетчаток . Выявлено четыре вида настроенных на диспаратность нейронов: возбуждающе настроенные на диспаратность (tuned-excitatory, TE), тормозно настроенные на диспаратность (tuned-inhibitory, TI), «дальние» (far, F) и «ближние» (near, N). TE нейроны отвечают максимальной частотой разрядов на стимулы с той диспаратностью, на которую они настроены. То есть при одновременной надлежащей стимуляции обоих глаз частота разрядов нейрона больше суммы двух монокулярных реакций. TI нейроны отвечают минимальной частотой разрядов на стимулы с предпочтительной диспаратностью. N нейроны отвечают максимальной частотой разряда на стимулы, находящиеся ближе точки фиксации, а F нейроны — на стимулы, находящиеся дальше точки фиксации. Среди N и F нейронов также выделяют настроенные на диспаратность нейроны. TI, N и F нейроны обычно обладают ярко выраженной глазодоминантностью — их ответ значительно зависит от изменения одного из двух монокулярных стимулов. Напротив, бинокулярные входы TE нейронов сбалансированы — их ответ в равной степени зависит от изменения любого монокулярного стимула.
У человека бинокулярны приблизительно 84 % нейронов зрительной коры . Процент и тип бинокулярных нейронов обезьяны ( макаки ) зависит от поля зрительной коры. В поле V1 бинокулярные нейроны составляют 65 %, из них 53 % — возбуждающе настроенные на диспаратность , 16 % — ближних, 19 % — дальних и 12 % — тормозно настроенных на диспаратность. В поле V2 бинокулярные нейроны составляют 78 %, из них 48 % — возбуждающе настроенные на диспаратность, 32 % — ближних, 15 % — дальних и 5 % — тормозно настроенных на диспаратность .
На случайно-точечные стереограммы реагируют только около 50 % бинокулярных нейронов коры.
Интересно отметить, что выделение монокулярно заданных контуров происходит в поле V1, в то время как выделение бинокулярно заданных контуров — только на следующей стадии переработки зрительной информации, в V2. Причём при фузии учитываются совпадения не между препарированными заранее контурами, а между точками или областями частей стереограммы.