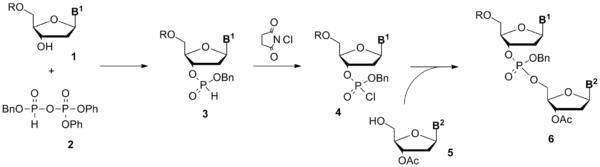
Синтез олигонуклеотидов
- 1 year ago
- 0
- 0

Эксцизио́нная репара́ция нуклеоти́дов ( англ. Nucleotide Excision Repair, NER ) — один из механизмов репарации ДНК . Наряду с эксцизионной репарацией оснований и репарацией ошибочно спаренных нуклеотидов , он позволяет исправить однонитевые повреждения ДНК , используя в качестве матрицы неповреждённую комплементарную цепь. В отличие от вышеуказанных механизмов, NER предназначен для более крупных повреждений ДНК, таких как пиримидиновые димеры , образующиеся в ДНК под действием ультрафиолета (УФ) .

У прокариот эксцизионная репарация нуклеотидов осуществляется системой белков Uvr . Три из этих белков — UvrA, UvrB и UvrC — образуют эндонуклеазу , известную как . Сначала белок UvrA распознаёт пиримидиновые димеры и прочие крупные повреждения и связывается с UvrB. Далее UvrA диссоциирует с затратой АТФ , а к UvrB присоединяется UvrC, который вносит надрезы в ДНК по обе стороны от повреждения: с отступом в 7 нуклеотидов от 5'-конца и с отступом в 3—4 нуклеотида от 3'-конца. Внесение надрезов требует затраты АТФ. Далее хеликаза UvrD расплетает ДНК между насечками, благодаря чему повреждённая цепь высвобождается. Синтез новой цепи взамен повреждённой осуществляет , хотя её могут заменять и III . В 99 % случаев при эксцизионной репарации, опосредованной системой Uvr, заменяется фрагмент ДНК длиной около 12 пар оснований (п. о.). В 1 % случаев заменяются более протяжённые участки — длиной около 1500 п. о., а в исключительных случаях и более 9000 п. о. Механизмы, регулирующие длину заменяемого фрагмента (короткий или длинный), неизвестны .
Комплекс Uvr может не только сам распознавать повреждения, но и направляться к ним другими белками. Так, если повреждение ДНК мешает транскрипции , белок смещает РНК-полимеразу и привлекает комплекс Uvr для устранения повреждения. Когда репарация матричной цепи ДНК завершается, транскрипция продолжается и образуется нормальный транскрипт .

У эукариот существует два механизма эксцизионной репарации нуклеотидов: репарация всего генома и репарация, связанная с транскрипцией. При первом пути белок узнаёт повреждение в любом месте генома. У млекопитающих белок XPC входит в состав комплекса, распознающего повреждение, в который также входят белки HR23B и . XPC также распознаёт повреждения, которые не может устранить эксцизионная репарация нуклеотидов, например, короткие участки частично денатурированной ДНК. Для распознавания некоторых типов повреждений, например, пиримидиновых димеров, XPC нуждается в дополнительных белках, которые помогают ему связаться с местом повреждения .
При втором пути, связанном с транскрипцией, повреждение распознаётся самой РНК-полимеразой II , при этом фермент останавливает движение по матрице ДНК. В некоторых случаях для протекания процесса фермент должен быть особым образом модифицирован или даже разрушен. Так, при остановке РНК-полимеразы II на месте пиримидинового димера её большая субъединица деградирует .
Собственно репарация в двух случаях осуществляется сходными наборами белков. В месте повреждения транскрипционный фактор , обладающий хеликазной активностью, раскручивает участок длиной около 20 п. о. Далее эндонуклеазы и вносят с обеих сторон надрезы в месте повреждения. Эндонуклеазы входят в состав комплекса , включающий также белок . Благодаря этому комплексу ERCC4 удерживается в связанном с ДНК виде в месте повреждений. Как правило, при эксцизионной репарации нуклеотидов у эукариот удаляется фрагмент длиной 25—30 п. о. Одноцепочечный повреждённый участок замещается за счёт синтеза новой цепи, который проводят и , а комплекс лигазы III и лигируют разрыв .
Если на пути репликативной вилки оказывается пиримидиновый димер, не удалённый системами репарации, то для дальнейшей репликации необходимо участие .
Мутации в разных белках, принимающих участие в эксцизионной репарации оснований, приводят к пигментной ксеродерме — аутосомно-рецессивному заболеванию, при котором солнечный свет и особенно УФ вызывают повреждения кожи , больные предрасположены к заболеванию раком . У больных синдромом Коккейна при остановке РНК-полимеразы II на месте УФ-повреждения не происходит деградации большой субъединицы. Это нарушение репарации приводит к неврологическим нарушениям и проблемам с ростом. Больные синдромом Коккейна, как и пациенты с пигментной ксеродермой, чувствительны к солнечному свету, но не предрасположены к развитию рака. Мутация одного из компонентов TFIIH, XPD, приводит к развитию трихотиодистрофии .
|
|
|
|---|
| Другие виды репарации | |
|---|---|
| Другие белки | |
| Регуляция | |




