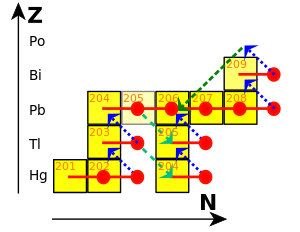
Мирные ядерные взрывы
- 1 year ago
- 0
- 0

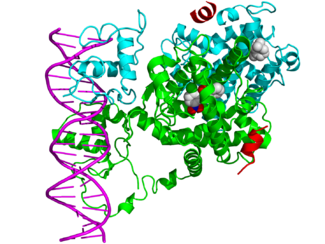
Ядерные поры , или ядерные поровые комплексы , — крупные белковые комплексы, пронизывающие ядерную мембрану и осуществляющие транспорт макромолекул между цитоплазмой и ядром клетки . Переход молекул из ядра в цитоплазму и в обратном направлении называется ядерно-цитоплазматическим транспортом .
Ядерные поровые комплексы ( англ. nuclear pore complex, NPC ) устроены сходным образом у всех исследованных на сегодняшний день организмов. Они образованы множеством копий примерно 30 разных белков-нуклеопоринов . Масса ядерных поровых комплексов колеблется в пределах от ~44 МДа в клетках дрожжей до ~125 МДа у позвоночных .
По данным электронной микроскопии [ источник не указан 967 дней ] , ядерные поры в поперечном сечении имеют форму «восьмиспицевого тележного колеса» [ источник не указан 967 дней ] , то есть имеют 8-лучевую симметрию. Эти данные подтверждает тот факт, что молекулы нуклеопоринов присутствуют в составе ядерной поры в количестве, кратном восьми. Проницаемый для молекул канал располагается в центре структуры. Ядерные поровые комплексы заякорены на ядерной оболочке с помощью трансмембранной части, от которой к просвету канала обращены структуры, получившие название спиц ( англ. spokes ), по аналогии со спицами тележного колеса. Эта коровая часть поры, построенная из восьми доменов , с цитоплазматической и ядерной сторон ограничена соответственно цитоплазматическим и ядерным кольцами ( англ. rings ; у низших эукариот они отсутствуют). К ядерному кольцу прикреплены белковые, направленные внутрь ядра, тяжи ( ядерные филаменты , англ. filaments ), к концам которых крепится терминальное кольцо ( англ. terminal ring ). Вся эта структура носит название ядерной корзины ( англ. nuclear basket ). К цитоплазматическому кольцу также прикреплены направленные в цитоплазму тяжи — цитоплазматические филаменты . В центре ядерной поры видна электрон-плотная частица, «втулка» или транспортёр ( англ. plug ).
Нуклепорины, белки, из которых построены ядерные поры, делят на три подгруппы. К первой относят трансмембранные белки, заякоривающие комплекс в ядерной оболочке. Нуклепорины второй группы содержат характерный аминокислотный мотив — несколько раз повторенные FG, FXFG или GLFG — последовательности (так называемые FG-повторы , где F — фенилаланин , G — глицин , L — лейцин , X — любая аминокислота). Функция FG-повторов, по-видимому, заключается в связывании , необходимых для осуществления ядерно-цитоплазматического транспорта. Белки третьей подгруппы не имеют ни мембранных доменов, ни FG-повторов, наиболее консервативны среди всех нуклеопоринов, их роль, по-видимому, заключается в обеспечении связывания FG-содержащих нуклепоринов с трансмембранными. Нуклеопорины также отличаются по своей мобильности в составе ядерной поры. Некоторые белки связаны с конкретной порой на протяжении всего клеточного цикла , в то время как другие полностью обновляются всего за несколько минут.
Количество ядерных пор на одно ядро может колебаться от 190 у дрожжей, 3000—5000 в клетках человека до 50 млн в зрелых ооцитах шпорцевой лягушки ( лат. Xenopus laevis ). Этот показатель может также варьировать в зависимости от типа клетки, гормонального статуса и стадии клеточного цикла . Например, в клетках позвоночных количество ядерных пор удваивается на протяжении S-фазы, одновременно с удвоением хромосом . При разборке ядерной оболочки во время митоза ядерные поры позвоночных распадаются на субкомплексы с массами около миллиона дальтон. Показано, что разборка ядерного порового комплекса инициируется циклин B-зависимой киназой , фосфорилирующей нуклеопорины. После завершения клеточного деления ядерные поры собираются de novo . Ядерные поры интерфазного ядра перемещаются большими массивами, а не независимо друг от друга, причём эти перемещения происходят синхронно с перемещениями ядерной ламины . Это служит доказательством того, что ядерные поры механически связаны между собой и формируют единую систему ( англ. NPC network ).