Мирные ядерные взрывы
- 1 year ago
- 0
- 0


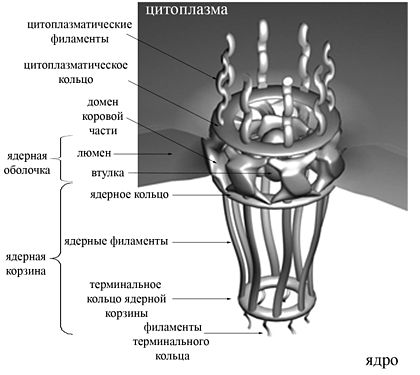
Я́дерные спе́клы , или спе́клы , или B-снурпосо́мы , или компа́ртменты фа́кторов спла́йсинга , или доме́ны SC-35 , или кла́стеры интерхромати́новых гра́нул ( англ. nuclear speckles ) — ядерные тельца , регулирующие сплайсинг . В ядерных спеклах находятся и малые ядерные рибонуклеопротеины . Изменения в белковом составе и функционировании ядерных спекл приводят к изменениям в альтернативном сплайсинге пре- мРНК , поэтому считается, что ключевой функцией ядерных спекл является регуляция доступности факторов сплайсинга в сайтах транскрипции . Недавние исследования показали, что в ядерных спеклах также находятся белки, задействованные в регуляции локализации хромосом , модификации хроматина , транскрипции, процессинге 3'-концов транскриптов, модификации мРНК, белки, полностью покрывающие мРНК, и мРНК-рибонуклеопротеины, поэтому ядерные спеклы рассматриваются как крупнейшие центры регуляции всех этапов экспрессии ядерных генов .
В интерфазных ядрах клеток человека содержится от 20 до 50 ядерных спекл, имеющих диаметр в несколько мкм . Одна ядерная спекла состоит из интерхроматиновых гранул диаметром 20—25 нм , которые соединяются фибриллами и формируют кластер — ядерную спеклу. Размер и форма ядерных спекл может значительно меняться и отличаться в клетках разных типов. На размер и форму спекл оказывают влияние множество факторов, в числе которых уровень АТФ в клетке, фосфорилирование /дефосфорилирование различных белков, транскрипция генов, активируемых стрессом, ремоделирование хроматина комплексом , а также транскрипция, опосредуемая РНК-полимеразой II .
Распределение белков в ядерных спеклах неравномерно. Так, локализуется в центре спекл, а длинная некодирующая РНК и малые ядерные РНК находятся на периферии. Показано, что размер спекл зависит от содержания РНК в них .
По биофизическим свойствам ядерные спеклы и нуклеоплазма не имеют значительных различий, однако ядерные спеклы слегка плотнее окружающей их нуклеоплазмы. Сборка и стабильность ядерных спекл зависит от взаимодействий их белковых компонентов. Многие белки спекл имеют очень гибкие участки низкой сложности (то есть не имеющие выраженной пространственной структуры), которые играют важную роль в белок-белковых взаимодействиях и . Они также играют важную роль в — ещё одном механизме, который обеспечивает целостность ядерных спекл. На разделение фаз на границе спекл и нуклеоплазмы влияют температура , pH , ионная сила и посттрансляционные модификации белков с участками низкой сложности. Цепочки повторяющихся аминокислот , например, пять или более последовательно соединённых остатков гистидина , служат сигналами локализации в ядерных спеклах .
В течение интерфазы ядерные спеклы очень стабильны. Разрушение ядерной оболочки в ходе митоза запускает разборку ядерных спекл, в результате которой белки спекл оказываются в цитоплазме . Там они собираются в так называемые митотические интерхроматиновые гранулы, которые можно видеть в метафазе , анафазе и телофазе . После того как в дочерней клетке восстановится ядерная оболочка, большинство факторов сплайсинга покидают митотические интерхроматиновые гранулы и в течение 10 минут перемещаются в ядро, однако некоторые факторы (например, SRSF2) могут оставаться в цитоплазме до G1-фазы . Когда начинается транскрипция, к новообразованным транскриптам привлекаются факторы сплайсинга и процессинга РНК, что приводит к нуклеации ядерных спекл вблизи сайтов активной транскрипции. Возможно, поведение ядерных спекл в клеточном цикле регулируется циклинами , потому что единственным неподвижным белком в интерфазных спеклах является циклин L1 .
Как и другие ядерные тельца, ядерные спеклы постоянно динамически обмениваются белковыми молекулами с нуклеоплазмой. Например, сплайсосомные рибонуклеопротеины собираются в тельцах Кахаля до перемещения в спеклы; кроме того, они регулируют созревание 3'-концов гистоновых транскриптов в тельцах гистоновых локусов . В ядерных спеклах можно обнаружить белки параспекл PSF и , и часто спеклы и параспеклы располагаются близко друг к другу. Белок Pat1b локализуется в ядерных спеклах и PML-тельцах . Однако функциональное значение столь тесных связей между ядерными тельцами ещё нужно установить .
В ядерных спеклах происходят многие посттрансляционные модификации белков, такие как фосфорилирование, метилирование , , убиквитинилирование и -илирование. В спеклах были обнаружены 31 протеинкиназа , и обратимое фосфорилирование играет важную роль в внутриклеточной локализации белков спекл, например, факторов сплайсинга, обогащённых аргинином и серином (SRSFs). Фосфорилирование и дефосфорилирование этих белков влияют на сплайсинг и сборку белков в спеклах .
Ядерные спеклы тесно связаны с сигнальными путями , включающими производные фосфоинозитола ?! (PIs). Многие белки спекл могут непосредственно с ними связываться, другие регулируются PIs косвенно через активность PI-зависимых белков, а именно, протеинкиназ и убиквитинлигаз . Существуют также прямые указания на то, что PIs сами локализуются в ядерных спеклах. Они могут оказывать значительное влияние на ядерные спеклы, поскольку регулируемые ими сигнальные белки чрезвычайно многофункциональны, как, например, некоторые протеинкиназы, локализующиеся в спеклах .
В ядерных спеклах были обнаружены некоторые белки цитоскелета . Перестройки цитоскелета находятся под контролем белков спекл, участвующих в сигнальных путях PI ( , , /ENIGMA и ) и (L-пластин, и ). Цитоскелетные белки могут регулировать не только сборку ядра, но и транскрипцию. Полимеризация актина имеет важное значение для ядерного транспорта и транскрипции, поскольку перестройки ядерных спекл при подавлении РНК-полимеразы II и привлечение РНК-полимеразы II к активированным промоторам зависят от полимеризации актина. В ядерных спеклах, кроме того, накапливается мономерный актин .
Значительная доля белков ядерных спекл подвергается ковалентному присоединению убиквитина или подобных ему белков, таких как SUMO1, и . При этом в спеклах обнаружены не были. Присоединение SUMO-1 служит типичным сигналом, привлекающим белки в ядерные спеклы .
В ядерных спеклах были обнаружены молекулы РНК, в том числе поли( А )-содержащие РНК и разнообразные некодирующие РНК . Накопление мРНК в спеклах зависит от ядерного экспорта, поскольку при отключении этого процесса в спеклах накапливается значительное количество мРНК. Некоторые исследования показали, что в ядерных спеклах может идти и сам сплайсинг, однако это утверждение требует дополнительной проверки. К некодирующим РНК, накапливающимся в спеклах, относятся малые ядерные РНК, 7SK РНК и длинная некодирующая РНК MALAT1 . Эта РНК стабилизируется не поли(А)-хвостом, а при помощи консервативной триспиральной структуры, и накапливается в ядре. Она взаимодействует с некоторыми SRSFs и малой ядерной РНК . MALAT1 влияет на альтернативный сплайсинг, регулируя фосфорилирование и распределение по ядру факторов сплайсинга. Однако было показано, что MALAT1 не является жизненно необходимой для нормального развития у мышей .
Более половины белков ядерных спекл задействованы в регуляции транскрипции и сплайсинга. Спеклы формируются в непосредственной близости от сайтов транскрипции, опосредуемой РНК-полимеразой II, и даже содержат несколько её субъединиц . Многие белки спекл регулируют элонгацию транскрипции, например, элонгин (фактор элонгации транскрипции SIII) и . В спеклах можно найти белки, регулирующие активность РНК-полимеразы II, и белки, вовлечённые в репарацию ДНК . Показана роль ядерных спекл в пространственной организации транскрипционно активного хроматина. Многие белки спекл задействованы в эпигенетической регуляции экспрессии генов, например, в спеклах были выявлены гистоны, гистонацетилтрансферазы , , деацетилазы и белок , ответственный за переход эухроматина в гетерохроматин .
Показано участие ядерных спекл в процессинге и экспорте мРНК. В частности, спеклы играют важную роль в выборе сайта альтернативного полиаденилирования . В ядерных спеклах происходит (m 6 A). Метилированный аденозин изменяет структуру молекулы РНК и влияет на её взаимодействие с белками, что оказывает важное влияние на метаболизм РНК. В ходе ранних этапов сплайсинга к сплайсосоме привлекается комплекс сращивания экзонов (EJC). Многие белки спекл взаимодействуют с EJC и ускоряют последующие этапы экспрессии генов, такие как сплайсинг соседних сайтов, ядерный экспорт мРНК и трансляция .
Изменения в структуре и функционировании ядерных спекл происходят при самых разнообразных заболеваниях. Некоторые из них, связанные непосредственно с мутациями в генах белков и РНК спекл, относительно редки. К их числу относят пигментный ретинит , и .
Возникновение рака связано с нарушениями в альтернативном сплайсинге, которые вызваны изменениями в экспрессии генов, кодирующих факторы сплайсинга, что часто происходит в результате морфологических изменений спекл. При многих видах рака наблюдается сверхэкспрессия белков спекл, таких как . Повышенного уровня достаточно для запуска развития опухоли .
Многие вирусы в ходе жизненного цикла используют белки клетки-хозяина, такие как белки сплайсинга и ядерного экспорта. Так, SRSFs и белки контролируют сплайсинг РНК ВИЧ и других вирусов, влияя на трансляцию вирусных белков. В свою очередь, при вирусной инфекции меняются локализация и уровень факторов сплайсинга, в частности, они удаляются из ядерных спекл. Вирусы также используют белки, отвечающие за процессинг 3'-конца мРНК. Любопытно, что на функционирование ядерных спекл влияют даже вирусы, реплицирующиеся в цитоплазме .
Ядерные спеклы вовлечены в патогенез многих , таких как болезнь Альцгеймера , болезнь Паркинсона , , боковой амиотрофический склероз , спинальная мышечная атрофия и шизофрения . Белки спекл вовлечены в дифференцировку нейронов и контролируют сплайсинг белков, специфичных для нейронов. Показана возможная роль ядерных спекл в заболеваниях, связанных с увеличением числа простых повторов в определённых генах .