Interested Article - Эволюция таламуса

- 2020-12-12
- 1
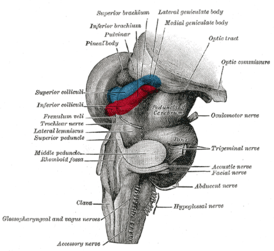
Эволюция таламуса — это процесс последовательного усложнения в ходе филогенеза хордовых структуры и функций таламуса . Эволюция таламуса у хордовых является одним из частных, но достаточно важных, проявлений общего процесса эволюции головного мозга , и, в целом, нервной системы у этих видов.
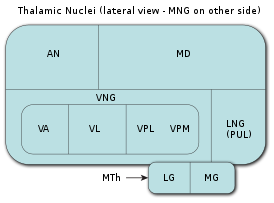
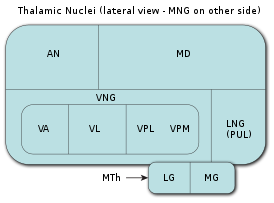
В таламусе выделяют эволюционно более древнюю часть — так называемый палеоталамус , , и эволюционно более молодую часть — неоталамус .
При сохранении общего эволюционно-консервативного плана строения таламуса, конкретные детали строения, степень сложности, общее количество ядер и функциональность таламуса очень сильно различаются у разных видов хордовых, стоящих на разных ступенях эволюционной лестницы.
Таламус анамниотических хордовых
Таламус у круглоротых рыб достаточно хорошо различим, в нём можно различить дорсальную и вентральную части («дорсальный таламус», или собственно таламус, и «вентральный таламус», или преталамус, субталамус). Однако в обеих частях таламуса у круглоротых всего по нескольку ядер. У костистых рыб таламус организован уже более сложно, в нём имеется несколько хорошо дифференцированных ядерных групп, он имеет больше связей с другими областями мозга рыбы. Большинство ядер таламуса костистых рыб занимаются обработкой соматосенсорной и зрительной информации, проекции их диффузны, слабо топографически локализованы. У земноводных таламус, особенно его эволюционно более молодая дорсальная часть («собственно таламус») уже значительно больше по размерам. Клеточный состав разных групп ядер таламуса земноводных сильнее различается, чем у рыб, но менее дифференцирован, чем у пресмыкающихся . Большая часть нейронов таламуса земноводных занимается обработкой зрительной информации, меньшая — соматической, слуховой или вестибулярной информации.
Таламус низших амниот
В таламусе пресмыкающихся уже можно найти чётко гомологичные ядрам таламуса млекопитающих группы ядер, с типичным именно для этих групп ядер гистологическим строением и типичным паттерном связей с другими частями мозга. У наиболее изученных модельных видов пресмыкающихся в дорсальном таламусе можно выделить 9 ядер, в вентральном таламусе (субталамусе или преталамусе) — 7 ядер. Афферентные и эфферентные связи таламуса с другими частями мозга у пресмыкающихся организованы значительно сложнее, чем у рыб и у земноводных. Латеральные (боковые) ядра таламуса пресмыкающихся принимают входящую информацию от ретиноталамического тракта (то есть от афферентных волокон сетчатки). Центромедиальные ядра таламуса пресмыкающихся занимаются обработкой и интеграцией зрительной и соматосенсорной информации, а вентрокаудальные — обработкой и интеграцией зрительной и слуховой информации, то есть те и другие занимаются мультисенсорной интеграцией и ассоциацией, что не свойственно таламусу рыб. У птиц таламус не намного сложнее таламуса пресмыкающихся: усложнению организации у птиц подверглась в большей степени контролирующая движения тела экстрапирамидная система , в частности стриатум и бледный шар (что, видимо, связано с необходимостью летать), чем таламус.
Таламус высших амниот (млекопитающих и человека)
Наибольшего развития таламус достигает у млекопитающих . У хищников он более развит, чем у грызунов или у травоядных животных, а наибольшего развития достигает у высших приматов и особенно у человека . Именно у млекопитающих дорсальный таламус («собственно таламус») стал основной релейно-ретрансляторной станцией, основным центром связи нижележащих отделов мозга с неокортексом , звеном, через который проходит и в котором ассоциируется, фильтруется и обрабатывается вся сенсорная, проприоцептивная, интероцептивная и моторная информация, за исключением информации от органов обоняния. Также именно у млекопитающих сформировалось множество замкнутых по принципу кольца двусторонних реципрокных связей таламуса с неокортексом, то есть таламо-кортико-таламических и кортико-таламо-кортикальных систем. Наиболее интенсивному развитию в ходе филогенеза млекопитающих подверглись так называемые ассоциативные ядра высшего порядка (или ассоциативные ядра верхнего уровня), занимающие дорсальную часть таламуса. В эти ядра поступает меньшее количество афферентных нервных волокон, чем в ассоциативные ядра низшего порядка, но они теснее связаны с ассоциативными областями коры. Именно эти ядра верхнего уровня, участвующие в образовании таламокортикальных ассоциативных систем, достигают наибольшего развития у высших приматов и человека. С работой именно этих ядер, наряду с работой неокортекса, связывают возникновение у приматов зачатков разума и самосознания.
Гомология таламуса хордовых и LAL членистоногих
В центральном нервном узле или головном мозге членистоногих обнаружена структура, гомологичная таламусу в головном мозге хордовых, как с точки зрения сходства процессов эмбрионального развития и паттернов экспрессии генов, так и с точки зрения сходства анатомического расположения в головном мозге, и с точки зрения сходства выполняемых физиологических функций (сбор информации и ретрансляция различных сенсорных путей в более передне расположенные части головного мозга или центрального нервного узла) — так называемые «боковые вспомогательные дольки» ( англ. lateral accessory lobes , LAL ).
Это заставляет предполагать, что, как минимум, зачаточными, примитивными структурами, гомологичными таламусу хордовых, или, по крайней мере, так называемым «эмбриональным потенциалом» для последующего в ходе эволюции раздельного, независимого развития у хордовых и у членистоногих подобных структур (то есть для проявления не гомологии, но гомоплазии) — то есть всем необходимым набором генов, которые у этого предка изначально могли выполнять какие-то другие функции, но затем в ходе эволюции были «рекрутированы» для сегментации у эмбриона промежуточных отделов мозга, паттернирования и специализации того отдела, который у хордовых известен как таламус, а у членистоногих — как LAL , — мог обладать уже гипотетический последний общий предок хордовых и членистоногих, или даже гипотетический последний общий предок всех билатерий (всех животных, обладающих билатеральной симметрией), — так называемая «урбилатерия». Это, в свою очередь, означает, что зачатки «прото-таламуса» в мозге эволюционных предков хордовых и членистоногих, или, как минимум, уже заложенный в имеющихся генах и в гибкости имеющейся программы эмбрионального развития мозга (её совместимости с таким «расширением» без возникновения сбоев в других частях программы развития эмбриона) эмбриональный потенциал для независимого появления таламуса у хордовых и LAL у членистоногих — появились между 550 и 600 млн лет назад. То есть таламус — по всей вероятности, структура эволюционно гораздо более древняя, чем было принято думать ранее.
Примечания
- The Medical Dictionary. (англ.) . The Medical Dictionary . FarLex Inc. Дата обращения: 28 сентября 2017. 18 сентября 2017 года.
- Merriam Webster Inc. (англ.) . Merriam Webster Medical Dictionary . Merriam Webster Inc.. Дата обращения: 28 сентября 2017. 18 сентября 2017 года.
- ↑ Edward G. Jones. : [ англ. ] : in 2 vol. . — редакция от 1985 года. — Нью-Йорк : Springer, 2012. — 915 с. — ISBN 978-1-4615-1749-8 . — doi : . — . — OCLC .
- ↑ Nicholas J. Strausfeld, Frank Hirth. : [ англ. ] : [ 3 октября 2017 ] // Science. — 2013. — Т. 340, № 6129 (12 April). — С. 157-161. — doi : . — PMID .
- ↑ Farries M.A. : [ англ. ] : [ 3 октября 2017 ] // Brain, Behavior and Evolution. — 2013. — Т. 82, № 4. — С. 211-214. — ISSN . — doi : . — PMID .
- 2020-12-12
- 1