Замедленное развитие
- 1 year ago
- 0
- 0

Эмбриональное развитие таламуса протекает в три основных этапа: формирование первичных доменов таламуса, образование среднедиэнцефалического организатора, и последующее созревание таламуса с формированием его ядерной и зональной организации.
Эмбриональный таламический комплекс состоит из периталамуса (или, иначе, преталамуса, ранее также называвшегося «вентральным таламусом»), среднедиэнцефалического организатора (который позже, в процессе эмбрионального развития таламуса, образует так называемый , и собственно таламуса (дорсального таламуса).
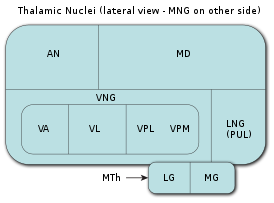
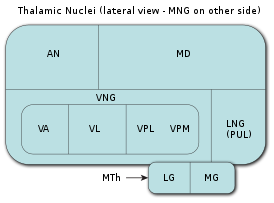
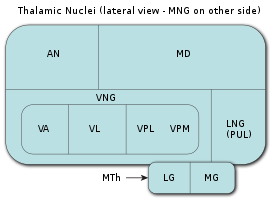
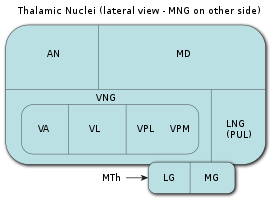
Таламус является самой крупной мозговой структурой, происходящей из эмбрионального диэнцефалона (зародышевого промежуточного мозга), расположенной между нижележащими структурами среднего мозга (мезэнцефалона) и вышележащими структурами большого мозга (церебрума), в частности, корой больших полушарий мозга.
У эмбриона человека уже на стадии Карнеги 9, то есть ещё до завершения нейруляции и формирования первичной нервной трубки , ещё на стадии загибания внутрь концов первичной нервной пластинки , в ней становятся различимы отдельные нейромеры , в том числе самый ростральный (самый передне расположенный) прозомер P, зачаток будущего переднего мозга (прозэнцефалона) . Позднее этот зачаток становится передним первичным мозговым пузырём (прозэнцефалоном) . Затем этот первичный мозговой пузырь делится на два вторичных мозговых пузыря — телэнцефалон (конечный мозг) и диэнцефалон (промежуточный мозг) . Ещё чуть позже в развивающемся промежуточном мозге (диэнцефалоне) эмбриона образуются два вторичных прозомера — D1 и D2. Из прозомера D2, собственно, и развиваются в дальнейшем таламус, а также эпиталамус и субталамус (преталамус), в то время как из прозомера D1 развивается в дальнейшем гипоталамус .
Данные, полученные в результате изучения процессов эмбрионального развития мозга у различных модельных позвоночных организмов, позволяют выдвинуть гипотезу о том, что для правильного развития эмбрионального таламического комплекса решающее значение имеет взаимодействие между двумя факторами транскрипции , Fez и Otx. Фактор транскрипции Fez селективно экспрессируется в процессе эмбрионального развития мозга клетками именно в области преталамуса, и функциональные эксперименты с выключением соответствующего гена показывают, что экспрессия белка Fez необходима для правильного развития преталамуса. Позади развивающегося под влиянием белка Fez преталамуса, области экспрессии белков Otx1 и Otx2 примыкают и упираются в область экспрессии белка Fez (то есть в будущий преталамус). Эти два белка, Otx1 и Otx2, необходимы для правильного развития таламуса.
В процессе раннего эмбрионального развития таламуса формируются два его первичных домена, каудальный домен (так называемый домен TH-C) и ростральный домен (так называемый домен TH-R). Каудальный домен эмбрионального таламуса служит источником клеток-предшественников для развития всех глутаматергических нейронов таламуса взрослых особей позвоночных животных, в то время как ростральный домен эмбрионального таламуса служит источником клеток-предшественников для развития всех ГАМКергических нейронов таламуса взрослых особей позвоночных животных.
В области стыка между доменами экспрессии белков факторов транскрипции Fez и Otx (то есть на границе между будущим преталамусом, и будущим таламусом), в эмбриональном таламическом комплексе формируется так называемая среднедиэнцефалическая организующая структура, или среднедиэнцефалический организатор. Среднедиэнцефалический организатор является своего рода «дирижёром оркестра», главным организатором всего последующего процесса эмбрионального развития таламуса и преталамуса, рассылающим необходимые для правильной дифференцировки клеток ядер таламуса и преталамуса межклеточные сигналы. Отсутствие среднедиэнцефалического организатора приводит к отсутствию таламуса и нередко также преталамуса в развивающемся мозге эмбриона. Сам среднедиэнцефалический организатор созревает в процессе эмбрионального развития таламического комплекса в направлении от более вентральных его частей, созревающих раньше, к более дорсальным, которые созревают позже. Белки, принадлежащие к семействам SHH и Wnt, являются главными регуляторными и дифференцировочными сигналами, испускаемыми средне-диэнцефалическим организатором.
Кроме своей функции как «дирижёра оркестра», управляющего всем процессом дальнейшего эмбрионального развития таламуса и преталамуса, среднедиэнцефалический организатор впоследствии созревает в особую гистологическую структуру внутри таламуса, так называемую ограниченную внутриталамическую зону.
Сразу после своего первоначального образования, среднедиэнцефалический организатор начинает выполнять свою роль главного дирижёра всего дальнейшего процесса эмбрионального развития таламуса и преталамуса из зачаточного таламического комплекса. Эту роль он выполняет, выделяя такие сигнальные молекулы, как SHH. У мышей и других млекопитающих, функциональную роль сигнальных молекул белка SHH, выделяемых среднедиэнцефалическим организатором, в дирижировании процессом дальнейшего эмбрионального развития таламуса и преталамуса из зачаточного таламического комплекса, непосредственно выяснить не удалось, поскольку генетическая мутация, приводящая к отсутствию функционального белка SHH, приводит к полному отсутствию у развивающегося зародыша даже не только зачатков таламического комплекса, но и всего диэнцефалона.
Тем не менее, исследования на развивающихся эмбрионах курицы показали, что экспрессия среднедиэнцефалическим организатором сигнального белка SHH является одновременно и необходимым, и достаточным условием для последующей индукции экспрессии генов, управляющих дифференцировкой клеток таламуса и преталамуса, и, соответственно, для их правильного развития. Исследования на другом модельном организме, рыбках данио-рерио, показали, что экспрессия двух генов семейства SHH, так называемых SHH-a и SHH-b (ранее также известного как twhh), определяет границы зоны среднедиэнцефалического организатора, и что сигнальные молекулы SHH необходимы и достаточны для начальной индукции молекулярной дифференцировки клеток будущего таламуса и преталамуса, но не являются обязательными для их дальнейшего поддержания и созревания. Кроме того, исследования на рыбках данио-рерио показали, что сигнальные молекулы SHH, исходящие из среднедиэнцефалического организатора, необходимы и достаточны для индукции дальнейшей дифференцировки и созревания как таламуса, так и преталамуса, в то время как сигналы SHH, исходящие из более вентральных по отношению к развивающимся таламусу и преталамусу областей мозга, большого значения для развития этих структур не имеют, и отсутствие вентрально исходящих сигналов SHH не приводит к нарушению развития таламуса и/или преталамуса, в отличие от сигналов SHH, исходящих от среднедиэнцефалического организатора.
Воздействие градиента экспрессии белка SHH, продуцируемого среднедиэнцефалическим организатором, приводит к дифференцировке нейронов будущего таламуса и преталамуса. Градиент экспрессии белка SHH, продуцируемого среднедиэнцефалическим организатором, вызывает формирование волны градиента экспрессии белка пронейрального гена нейрогенина-1, распространяющейся в направлении от задней части к передней, в основном (каудальном) домене таламического зачатка, и одновременно — формирование волны градиента экспрессии белка Ascl1 (ранее известного как Mash1) в оставшейся узкой полоске рострально расположенных клеток таламического зачатка, непосредственно прилегающей к среднедиэнцефалическому организатору (то есть в ростральном домене таламического зачатка), и в преталамусе.
Формирование этих специфических зональных градиентов экспрессии тех или иных пронейральных белков приводит к дальнейшей дифференцировке глутаматергических «релейно-ретрансляторных» нейронов из расположенных в каудальном домене таламического зачатка нейрогенин-1-положительных клеток-предшественников, и к дифференцировке ГАМКергических ингибирующих нейронов из расположенных в ростральном домене таламического зачатка, непосредственно прилегающем к средне-диэнцефалическому организатору, и в преталамусе Ascl1-положительных клеток-предшественников. У рыб выбор одного из этих двух альтернативных путей дифференцировки для каждой конкретной клетки-предшественника в той или иной зоне зачаточного таламического комплекса управляется динамической экспрессией белка Her6, являющегося гомологом белка HES1 человека. Экспрессия этого фактора транскрипции, относящегося к семейству «волосообразных» белков bHLH, приводит к подавлению экспрессии гена нейрогенина-1, однако необходима для поддержания и усиления экспрессии белка Ascl1. В процессе дальнейшего эмбрионального развития таламического зачатка экспрессия белка Her6 и, соответственно, связанное с ним подавление экспрессии белка нейрогенина-1 и усиление экспрессии белка Ascl1 постепенно исчезает в каудальном домене таламического зачатка, в то время как в преталамусе и в узкой полоске рострально расположенных таламических клеток, примыкающей к среднедиэнцефалическому организатору, экспрессия белка Her6 и, соответственно, подавление экспрессии белка нейрогенина-1 и усиление экспрессии Ascl1 усиливается и нарастает. Это делает каудально-ростральный градиент экспрессии нейрогенина-1/Ascl1 более выраженным, границы доменов более чёткими, и способствует завершению созревания и дифференцировки клеток таламуса и преталамуса. Исследования на развивающихся эмбрионах курицы и мыши показали, что блокада сигнального пути белка SHH в этот период эмбрионального развития приводит к полному отсутствию рострального домена таламического зачатка и к значительному уменьшению размеров каудального домена таламического зачатка. Ростральный домен таламического зачатка даёт начало ГАМКергическим ингибирующим нейронам таламуса, расположенным в основном в ретикулярном ядре таламуса взрослых животных, в то время как каудальный домен таламического зачатка даёт начало глутаматергическим «релейно-ретрансляторным» нейронам, составляющим основную часть клеток таламуса, и подвергающимся дальнейшей дифференцировке с образованием отдельных таламических ядер и групп ядер.
Было показано, что у человека часто встречающаяся генетическая вариация в области промотора гена белка транспортера серотонина (SERT), а именно, обладание длинной (SERT-long) или короткой (SERT-short) аллелями этого гена (гена 5-HTTLPR), влияет как на эмбриональное, так и на последующее (постэмбриональное) развитие и созревание определённых областей таламуса и на их конечный размер у взрослых. Люди, у которых имеется две «коротких» аллели гена 5-HTTLPR (SERT-ss), имеют больше нейронов в ядрах подушки таламуса и более крупный размер этих ядер, а также, возможно, больше нейронов и более крупный размер лимбических ядер таламуса (ядер, поддерживающих связь с эмоциональными центрами лимбической системы), по сравнению с гетерозиготами по этому гену или обладателями двух «длинных» аллелей гена 5-HTTLPR. Увеличение размеров этих структур таламуса у таких людей предполагается в качестве части анатомического объяснения того, почему люди, у которых имеется две «коротких» аллели гена 5-HTTLPR, в большей мере, чем люди, гетерозиготные по этому гену или обладающие двумя «длинными» аллелями гена 5-HTTLPR, предрасположены к таким психическим расстройствам , как большое депрессивное расстройство , посттравматическое стрессовое расстройство (ПТСР), а также к суицидальным тенденциям и попыткам .