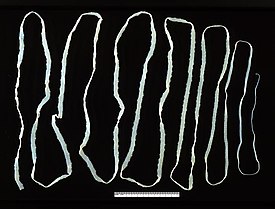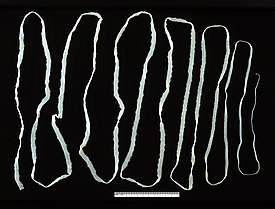
Бычий форум
- 1 year ago
- 0
- 0

Бы́чий о́вод , или обыкнове́нный подко́жник , или строка́ ( англ. Hypoderma bovis ) , — вид паразитических двукрылых из семейства оводов (Oestridae), широко распространённый в Северном полушарии . На личиночных стадиях паразитирует на крупном рогатом скоте , изредка на других копытных ( лошадях , овцах ) и на человеке, вызывая заболевание гиподерматоз ; во взрослом состоянии не питается . Ввиду причинения значительного урона животноводству был целенаправленно истреблён в ряде стран Европы во второй половине XX века .
Вид практически повсеместно распространён в Голарктике в умеренной зоне , заходя в субтропики , — на территории Евразии (кроме севера Сибири ), Северной Африки и Северной Америки . Взрослые особи — теплолюбивые насекомые, предпочитающие низкую влажность. Куколки длительное время проводят в почве , также плохо перенося высокую влажность; они не выживают при затоплении, поэтому бычьи оводы редки в долинах крупных рек, время от времени выходящих из берегов , таких как Нил и Ред-Ривер . Значительные трудности для взрослых насекомых создаёт сильный ветер, из-за чего они не встречаются на открытых островах .
Довольно крупные мухи длиной 13—15 мм. Тело густо покрыто длинными волосками, которые придают насекомым характерную полосатую жёлто-чёрную окраску. Фасеточные глаза широко расставлены. Лоб составляет от трети до половины ширины головы (у самцов и самок соответственно), несёт 3 простых выпуклых глазка. Усик состоит из 3 члеников и ; второй членик укрупнён, частично скрывает третий. Ротовой аппарат полностью редуцирован (взрослая особь не питается) .
Грудь хорошо выражена; как и у других двукрылых, наиболее мощно развита среднегрудь . Среднеспинка разделена поперечным швом и окрашена в чёрный цвет, который в передней половине замаскирован густыми жёлтыми волосками. На задней части среднеспинки волоски чёрные; они отсутствуют на 4 продольных голых полосах. Мезоплевры покрыты жёлтыми волосками, по пучку таких же волосков расположено на трёх других боковых склеритах (птероплеврах, стерноплеврах и гипоплеврах), остальные боковые щетинки чёрного цвета. Задние ноги несколько длиннее передних, по длине примерно равны телу; у них утолщены основания бёдер и средняя часть голеней. Лапка 5-члениковая, с более крупным первым члеником (на задних ногах его длина примерно равна четырём другим членикам лапки, взятым вместе). Пульвиллы развиты хорошо, по размерам близки коготкам. Крылья в длину составляют 9—11 мм, мембрана прозрачно-коричневатая, жилки коричневые. В спокойном состоянии крылья складывают не полностью, их концы немного раздвинуты .
Брюшко составлено 5 основными сегментами, а также ещё несколькими, которые преобразованы в копулятивный орган (фаллосому) у самцов и яйцеклад у самок. Расположение волосков на брюшке также формирует полосы: на II, IV и V тергитах — жёлтые (на переднем — более светлые, на задних — рыжего оттенка), на III тергите они чёрные. Втяжной яйцеклад самок имеет телескопическое строение; он подвижный и длинный (в выдвинутом состоянии — до 11 мм). Большую его часть составляют VI и VII сегменты брюшка, на них сохраняются небольшие тергиты и стерниты, разделённые мембранами .

Яйца овальной формы, длиной около 1 мм, с ячеистым хорионом . Микропилярное отверстие сохраняется в виде небольшого углубления в хорионе. Самки откладывают яйца по отдельности, присоединяя к волосам жертвы с помощью придатка яйца. Прикрепительный придаток соединён к яйцом коротким стебельком и имеет форму свёрнутой вокруг волоса пластинки (расправляется в концентрированной щелочи ). Предположительно, адгезия достигается либо исключительно за счёт деформации придатка при высыхании после откладки, либо также при участии клейких выделений придаточных желёз яйцеклада. Видимой крышечки ( лат. operculum ) у яйца нет; перед выходом личинка вскрывает незаметный снаружи дуговидный шов, проходящий в передней части яйца ниже микропиле .
Личинки бычьих оводов имеют типичное для круглошовных мух строение и переходят на стадию куколки после третьей линьки. Они лишены ног и состоят из ввёрнутой в грудные сегменты головы , а также видных снаружи (видоизменения шейного сочленения), 3 грудных и 8 брюшных сегментов. На переднем конце тела находится вторичное ротовое отверстие, ведущее в предротовую полость. На переднем крае большинства сегментов с брюшной стороны находятся поля коротких жёстких шипиков, участвующих в движении; у личинок II и III возраста к ним добавляются поля шипиков на заднем крае сегментов .
Личинки I возраста подвижны. Они характеризуются светлоокрашенными, сравнительно мягкими покровами, делающими возможным значительный межлиночный рост: от 0,6 мм длины при выхода из яйца до 17 мм перед первой линькой (объём тела увеличивается более чем на три порядка). В начале развития личинки обладают веретеновидной формой, по мере миграции, роста и достижения окончательной локализации в хозяине становятся широкоовальными. Из-за растяжения кутикулы поля шипиков становятся крайне разреженными и скоро теряют функциональное значение для передвижения. Вероятно, на этом этапе ключевую роль в прокладывании хода в тканях хозяина приобретают выделяемые личинкой гидролитические ферменты :825 . В предротовой полости лежит пара сравнительно крупных ротовых крючьев ( лат. hamuli ) . Стигмы представлены лишь одной парой на последнем сегменте .
Личинки II возраста малоподвижны, вооружение из шипиков у них развито слабее, чем у личинок III возраста. На границе между I и II грудными сегментами появляется пара передних стигм, а задняя пара значительно укрупняется (состоит из нескольких десятков пор). Ротовой аппарат значительно редуцируется: крючья преобразуются в небольшие неподвижные склериты, но глоточный отдел остаётся хорошо развитым .
Личинка III возраста отличается сильно вздутой брюшной стороной тела, так что последний сегмент брюшка заметно загнут на спину. Задняя пара стигм приобретает ещё большие размеры, добавляется ещё по одной паре рудиментарных стигм на II—VI брюшных сегментах. Кутикула личинки III возраста (предкуколки) очень рельефна, границы сегментов чёткие, по бокам тела появляется пара продольных борозд. На ней уже виден пупарный шов, намечающий положение крышечки ( лат. operculum ) для вылета взрослого насекомого: он образует замкнутый контур на спинной стороне между I грудным до I брюшным сегментами. Окраска серая или коричневая .
При окукливании личинка несколько сжимается, не выходя из старой кутикулы, куколка , таким образом, развивается внутри пупария , или ложного кокона. Взрослое насекомое покидает пупарий, отбрасывая крышечку .
Анатомия изучена преимущественно на личиночных стадиях. Пищеварительная трубка подразделена двумя клапанами на три отдела: пищевод, среднюю кишку (состоит из провентрикулуса, собственно средней кишки и короткой тонкой кишки) и заднюю кишку. От задней кишки отходят 4 мальпигиевых сосуда . Дыхательная система представлена двумя продольными трахейными стволами, которые разветвляются в сегментах тела. Нервная система личинок включает два крупных ганглия — надглоточный и грудной, а также несколько небольших трахейных ганглиев (в передней части трахейных стволов). У имаго к ним добавляется самостоятельный подглоточный ганглий .
Формирование половой системы и гаметогенез полностью завершаются на стадии куколки. У самцов развивается пара сферических семенников , которые связанны общим протоком с фаллосомой. У самок имеется пара яичников , протоки которых открываются на яйцекладе и соединены с придаточными железами и 3 семяприёмниками .
Определение пола особей хромосомное, как и у многих других представителей двукрылых, осуществляется по схеме XY. Нормальный кариотип самок — 12, XX , у самцов — 12, XY (n=6). Половые хромосомы самые крупные (X — метацентрическая, Y — примерно в полтора раза меньше, субтелоцентрическая) .
Время лёта взрослых насекомых приурочено к летним месяцам и чётко синхронизировано в популяции по внешним условиям. Короткоживущие имаго вылетают в утренние часы и в массе собираются в характерных биотопах, не меняющихся из года в год. В таких местах одномоментное количество самцов всегда значительно выше, поскольку самки вскоре после спаривания (в отличие от самцов) отправляются на поиски крупного рогатого скота для откладки многочисленных (до 600—800) яиц на шерсть животных (как правило, на ноги). Каждое отдельное яйцо фиксируется самкой на волосе прикрепительным придатком. Обычно через 3—5 дней после вылета взрослые особи гибнут, но в условиях плохой погоды из-за низкой физиологической активности продолжительность их существования может увеличиваться до месяца .
Вылупление миниатюрной (0,6 мм) безногой личинки I возраста происходит через 3—7 дней после яйцекладки. Она спускается по волосу и вбуравливается в кожу хозяина, после чего наступает продолжительная фаза миграции по соединительной ткани, длящаяся от 4 до 8 месяцев. Сравнительно редко личинок бычьего овода отмечают в стенках пищевода (в большинстве случаев это оказывается результатом ошибочного определения личинок I возраста другого вида оводов — , или пищеводника). По-видимому, обязательным этапом развития оказывается проникновение в спинномозговой канал , где личинки локализуются в эпидуральном жире. Осенью или зимой (в зависимости от климатических условий) они покидают спинной мозг хозяина и перемещаются под кожу спины животного, как правило, в межрёберные пространства не далее 25 см от хребта. Там личинка I возраста, к этому моменту достигшая за счёт межлиночного роста длины 17 мм, претерпевает первую линьку .
Под кожей хозяина личинки II возраста переходят к аэробному метаболизму : присутствие личинки вызывает образование и вскрытие фурункула , так что она разворачивается в своём ходе на 180° и выставляет наружу задний конец тела с парой крупных стигм . Питаются тканевым экссудатом , накапливая запасы жира (до трети массы). Выделения личинок обладает бактерицидной активностью, таким образом паразит препятствует развитию нагноения в фурункуле. Через 40—80 дней происходит линька на стадию личинки III возраста (предкуколку). Через некоторое время она выпадает наружу через разрыв кожи и перемещается в укрытие, где линяет на стадию покоящейся куколки. При этом кутикула личинки III возраста не сбрасывается полностью, а образует наружную оболочку куколки — пупарий . Куколки способны переносить короткие периоды отрицательных температур . Через 15—65 дней (в зависимости от времени выхода из хозяина и фенологических условий) происходит синхронный вылет взрослых насекомых с полностью развитой половой системой .
По-видимому, из-за того, что во внешней среде оказываются только короткоживущие взрослые насекомые и быстро прячущиеся в почву личинки III возраста и куколки, у бычьих оводов нет серьёзных естественных врагов. Отмечены отдельные случаи выклёвывания личинок из кожи коров галками и скворцами, а также эпизоды поедания взрослых особей хищными насекомыми .
Описаны несколько случаев паразитирования на куколках бычьих оводов других насекомых: наездников из рода Trichopria (семейство диаприид ) и мух из рода (семейство Fanniidae ). Очевидно, в обоих случаях имел место не гиперпаразитизм , а заражение свободных личинок III возраста до окукливания. Личинки как наездников, так и мух в описанных случаях не смогли завершить развитие, что может указывать на факт заражения неспецифического хозяина .
В области распространения бычий овод наносит серьёзный ущерб животноводству . Стаи самок, преследующие крупный рогатый скот, вызывают у животных реакцию панического бегства (так называемый «бзык», или «зык»). Бег сильно утомляет организм, может приводить к травмам конечностей, нарушает режим выпаса и заметно снижает удои в период лёта взрослых насекомых . Традиционно ошибочно считалось, что такое поведение коров связано с болевыми ощущениями, которые испытывают животные при укусах оводов или при внедрении яиц под кожу. Тем не менее давно установлено, что взрослые самки оводов не питаются, а яйца откладывают на волосяной покров. Реакция у телят носит врождённый характер и не требует обучения. Замечено, что в ходе бегства коровы стремятся к водоёмам, которых подкожники, напротив, избегают. Была высказана гипотеза, что это явление обусловлено неспособностью самок успешно прикрепить яйцо к волосу в условиях высокой влажности .
Заражённые личинками особи демонстрируют снижение удоя (на 10—15 %, при интенсивном заражении — на 25 %) и привеса (на 0,07—0,6 кг/день). Также в результате миграции и подкожного развития личинок ухудшается качество туши: вокруг ходов личинок и фурункулов формируется бесцветная желеобразная соединительная ткань ( англ. butcher jelly ) , снижающая товарный вид и пищевую ценность мяса (перерождение также затрагивает прилежащую мускулатуру) . Кроме того, при подкожной локализации личинки повреждают наиболее ценные для выделки части шкуры . Гибель личинки в теле хозяина может вызывать серьёзные воспалительные процессы (в том числе поражение спинного мозга), иногда с летальным исходом. По-видимому, такая реакция опосредована токсическим или аллергическим действием. Более старые особи демонстрируют низкое заражение подкожниками, вероятно, в результате формирования приобретённого иммунитета против личинок :2523 .
В редких случаях самки бычьего овода откладывают яйца на волосы человека. В случае успешного заражения характер миграции и конечная локализация личинки несколько меняются. Подкожные стадии в теле человека приурочены к коже верхней части спины, шеи и головы. Иногда личинки проникают в глаза, вызывают осложнения в спинном и даже головном мозге .
Наиболее простые методы защиты домашнего скота во время лёта бычьего овода состоят в размещении стада в дневные часы под образующими тень навесами и ночной выпас, также действенен загон стада в водоём, поскольку насекомые активны только днём и избегают тенистых и влажных биотопов. Подкожных личинок II и III возраста можно сравнительно легко извлечь хирургическим путём .
В первой половине XX века для сокращения численности подкожных оводов широко использовался «деррисовый порошок» — природный инсектицид ротенон , синтезируемый некоторыми видами бобовых и коровяком . В послевоенные годы его вытеснили синтетические фосфорорганические соединения . Наконец с 1980-х годов в качестве эффективной меры профилактики и лечения гиподерматоза стали применять подкожные инъекции ивермектина ; с конца 1990-х также вошёл в обиход :1343 .
В качестве ещё одной меры защиты были разработаны вакцины , одновременно действующие против бычьего овода и близких видов (например, ); тем не менее широкого применения вакцинация так и не получила. Мишенью для приобретённого иммунитета становится личинка I возраста, на поверхности которой имеются доступные антигены — белки гиподермины ( сериновые протеазы , обеспечивающие продвижение мигрирующей личинки по соединительной ткани хозяина) :825 .
Бычьи оводы обладают довольно узкой специфичностью к животным-хозяевам и годичным жизненным циклом, в связи с этим их популяции, несмотря на высокую плодовитость, оказываются уязвимыми на стадии зимующих личинок. Одна из первых успешных попыток истребления подкожных оводов была предпринята на рубеже XIX и XX века в островных фермерских хозяйствах Дании , метод состоял в хирургическом удалении подкожных личинок у всего поголовья и последующем санитарно-эпидемиологическом контроле импорта животных .
После начала интенсивного применения в зимний сезон различных инсектицидов ( ротенона , ДДТ , фосфорорганики и в особенности антибиотиков авермектинового ряда ) введение национальных и региональных программ позволило полностью истребить популяции на территории Дании , Нидерландов , Ирландии , Великобритании и Чехословакии , а также в ряде регионов Германии и Франции . Популяция подкожных оводов на территории Словакии впоследствии восстановилась .
Представления о родственных отношения бычьих оводов с другими представителями рода Hypoderma значительно варьируют. По данным молекулярной филогенетики, полученным разными исследователями, в различных комбинация их сближали с паразитами крупного рогатого скота ( и ) и паразитами оленей ( , и Hypoderma tarandi ) . По данным анализа 2016 года, который основывался на последовательности гена цитохром c-оксидазы (прежде всего, китайских популяций), подкожники крупного рогатого скота образуют монофилетическую группу и дивергировали от подкожников оленей около 4,5 млн лет назад (на рубеже миоцена и плиоцена ) .