Проклятие золотого цветка
- 1 year ago
- 0
- 0

Цветок — это видоизменённый и укороченный побег , формирующийся из генеративной меристемы . Органы, составляющие зрелый цветок, располагаются кругами: снаружи круг из чашелистиков , затем из лепестков , тычинок и в центре — из плодолистиков, образующих пестики . Считается, что они являются видоизменёнными листьями или выростами стебля . Эту идею впервые высказал И. В. Гёте в XVIII веке , назвав цветы «изменёнными листьями». Подобная точка зрения подтверждается результатами исследований гомеозисных мутаций . Изучение гомеозиса привело к формулировке модели формирования цветка ABC .
В процессе развития генеративного побега (как и при эмбриональном развитии животных) имеет значение позиционная информация, которая определяет направление дифференцировки и взаимное расположение тех или иных структур в процессе их закладки. По мере формирования цветка подобная разметка (spatial pattern formation) определяет взаимное расположение органов относительно друг друга, положение отдельных клеток внутри развивающихся тканей, распределение митотической активности и формирование в пространстве «образа» (pattern) цветка в целом. При флоральном органогенезе вначале происходит разметка положений органов, а только после этого начинается их закладка, которая у различных растений происходит по разным сценариям. После этого развитие контролируется совокупностью генов, описываемых моделью АВС.
Аксиоматические принципы разметки:
Независимо от этого цветок характеризуется мерностью (например, димерный, тримерный и т.п. околоцветник) и полнотой структуры цветка (числом тычинок, плодолистиков и элементов околоцветника). Мерность различных кругов может увеличиваться или уменьшаться в результате различных мутаций. При этом в одном цветке число элементов околоцветника может увеличиваться, а число тычинок и плодолистиков - уменьшаться, и наоборот. Существует корреляция между числом и положением тычинок и плодолистиков , а также между числом и расположением чашелистиков и лепестков . Полнота связана с размерами зон, внутри которых происходит закладка примордиев листочков околоцветника, тычинок или плодолистиков. Существует предположение, что у цветковых разметка биполярна и идет в двух направлениях: акропетальном и бизипетальном. При превращении вегетативной меристемы с акропетальной разметкой в генеративную её дифференцировка на центральную и периферическую зоны утрачивается. Это приводит к терминализации роста и к началу базипетальной разметки. Если формирование зоны базипетальной разметки отсрочено во времени, то структура цветка обогащается дополнительными элементами. Когда эта зона не формируется, цветок имеет максимальное (неопределенное) число органов. Терминализация роста связана с активацией генов класса С , и при нарушении их работы у мутантов развивается недетерминированный цветок. Если зона базипетальной разметки формируется ускоренно, часть органов не развивается, появляется неполный цветок с делецией органов. Сокращение общего времени разметки, вероятно, связано с нарушениями работы генов класса А . Оно приводит к исчезновению структур, которые размечаются последними; органы, размечающиеся первыми - самые стабильные. По стабильности органов можно судить о порядке их разметки.
У арабидопсиса, например, разметка идет так: 1. абаксиальный и адаксиальный чашелистики; 2. плодолистики; 3. длинные тычинки и лепестки; 4. короткие тычинки.
В соответствии с АВС-моделью генетической регуляции, существует предположение, что зона акропетальной разметки совпадает с зоной экспрессии генов класса А (разметка чашечки и венчика), а зона базипетальной разметки - с генами класса С (разметка гинецея и андроцея). Если рассматривать мутантные цветки арабидопсиса, то у махровой формы agamous (при вставке новых кругов органов, при которой утрачивается гинецей), или у мутанта apetala 2 (с делецией кругов) не только нарушается программа развития определенных органов, но и геометрия цветка меняется, то есть эта формальная замена одного органа другим не является гомеозисом в узком смысле этого термина. .

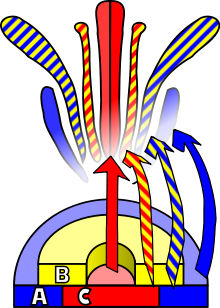
Так называемая АВС модель развития цветка - это современная парадигма биологии развития. Согласно этой модели дифференциация органов цветка определяется работой 3 классов регулирующих генов: гены класса А отвечают за развитие чашелистиков, в сумме с генами класса В они определяют формирование лепестков, совместная работа генов класса В и С ведет к развитию тычинок, а гены С сами по себе контролируют появление пестика в центре цветка. Эти гены кодируют факторы транскрипции, которые вызывают специализацию тканей растения в процессе развития.
Впоследствии были добавлены еще два класса генов: гены класса D, которые отвечают за развитие завязи в цветке, мутации по этому гену приводят к развитию плодолистиков вместо завязи, а сверхэкспрессия этих генов - к формированию завязей вместо чашелистиков и лепестков; и гены класса E, которые контролируют идентичность трех внутренних кругов.
При нарушении работы этих генов одни части цветка превращаются в другие (тычинки в лепестки или лепестки в чашелистики). Модельным видом в этих исследованиях был арабидопсис , у которого был обнаружен ряд гомеозисных мутаций, при совместном действии некоторых из которых все части цветка превращались в листья.
Модельный вид, на котором изучается цветение, – арабидопсис – является растением длинного дня. Однако, несмотря на то, что обычно он зацветает, когда световой день удлиняется, его можно «перепрограммировать» на короткий день. В процессе экспериментов было установлено, что изменения сроков зацветания арабидопсиса были обусловлены нарушениями в работе фоторецепторов. Инактивация криптохрома 2 блокирует процесс развития цветка, начинающийся при увеличении продолжительности дня, поэтому он, вероятно, выступает в качестве одного из стимуляторов цветения. Отсутствие фитохрома В, наоборот, ускоряет цветение, поэтому он, скорее всего, относится к ингибиторам этого процесса. Роль фоторецепторов в развитии растений подробно описана в данной публикации [см. “Fitocromos y desarrollo vegetal”, J.F.Martínez et.al.; Investigación y ciencia, febrero de 2002].
Из всех мутаций, связанных с цветением арабидопсиса, самой заметной является мутация гена Constans (CO), потому что она делает растение невосприимчивым к изменению длины светового дня. Идентификация гена СО быстро привела к выделению кодируемого им нового белка (СО), характерного только для растений, структура которого сходна со структурой активаторов транскрипции генов животных.