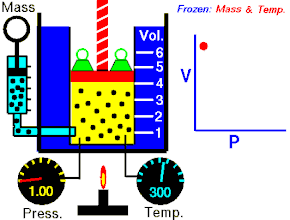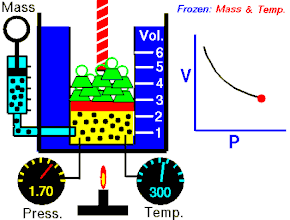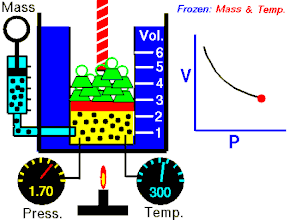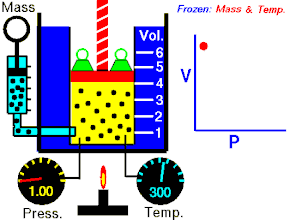
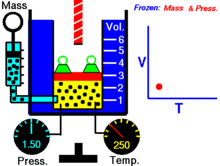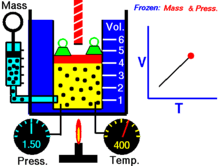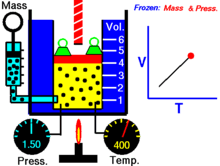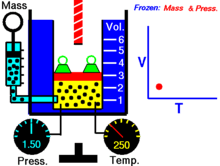
Закон Гука
- 1 year ago
- 0
- 0

Биогенетический закон Геккеля — Мюллера (также известен под названиями «закон Геккеля», «основной биогенетический закон», «теория эмбриональной рекапитуляции»): каждое живое существо в своём индивидуальном развитии ( онтогенезе ) повторяет, в известной степени, формы, пройденные его предками или его видом ( филогенез ) .

Биогенетический закон сыграл заметную роль в истории развития науки, однако в XX веке был опровергнут и не признается современной биологической наукой (см. далее ).
Фактически «биогенетический закон» был сформулирован ещё задолго до возникновения дарвинизма.
Немецкий анатом и эмбриолог Мартин Ратке (1793—1860) в 1825 г. описал жаберные щели и дуги у эмбрионов млекопитающих и птиц — один из наиболее ярких примеров рекапитуляции.
В 1824—1826 годах сформулировал «закон параллелизма Меккеля-Серра»: каждый организм в своём эмбриональном развитии повторяет взрослые формы более примитивных животных [ источник не указан 4527 дней ] .
В 1828 году Карл Максимович Бэр , основываясь на данных Ратке и на результатах собственных исследований развития позвоночных, сформулировал закон зародышевого сходства : «Эмбрионы последовательно переходят в своём развитии от общих признаков типа ко все более специальным признакам. Позднее всего развиваются признаки, указывающие на принадлежность эмбриона к определенному роду, виду, и, наконец, развитие завершается появлением характерных особенностей данной особи». Бэр не придавал этому «закону» эволюционного смысла (он до конца жизни так и не принял эволюционного учения Дарвина), однако позднее этот закон стал рассматриваться как «эмбриологическое доказательство эволюции» (см. Макроэволюция ) и свидетельство происхождения животных одного типа от общего предка.
«Биогенетический закон» как следствие эволюционного развития организмов впервые был сформулирован (довольно нечётко) английским естествоиспытателем Чарльзом Дарвином в его книге «Происхождение видов» в 1859 г: «Интерес эмбриологии значительно повысится, если мы будем видеть в зародыше более или менее затененный образ общего прародителя, во взрослом или личиночном его состоянии, всех членов одного и того же большого класса» (Дарвин Ч. Соч. М.-Л., 1939, т. 3, с. 636.)
За 2 года до формулировки Эрнстом Геккелем биогенетического закона сходную формулировку предложил на основе своих исследований развития ракообразных работавший в Бразилии немецкий зоолог Фриц Мюллер . В своей книге «За Дарвина» (Für Darwin), изданной в 1864 году, он выделяет курсивом мысль: «историческое развитие вида будет отражаться в истории его индивидуального развития».
Краткая афористичная формулировка этого закона была дана в 1866 году немецким естествоиспытателем Эрнстом Геккелем. Краткая формулировка закона звучит следующим образом: Онтогенез есть рекапитуляция филогенеза (во многих переводах — «Онтогенез есть быстрое и краткое повторение филогенеза») .
Уже в XIX веке было известно достаточно фактов, противоречащих биогенетическому закону. Так, были известны многочисленные примеры неотении , при которой в ходе эволюции происходит укорочение онтогенеза и выпадение его конечных стадий. В случае неотении взрослая стадия вида-потомка напоминает личиночную стадию вида-предка, а не наоборот, как этого следовало бы ожидать при полной рекапитуляции.
Также было хорошо известно, что, вопреки «закону зародышевого сходства» и «биогенетическому закону», весьма резко различаются по строению самые ранние стадии развития зародышей позвоночных — бластула и гаструла — и лишь на более поздних стадиях развития наблюдается «узел сходства» — стадия, на которой закладывается план строения, характерный для позвоночных, и зародыши всех классов действительно похожи друг на друга. Различия ранних стадий связаны с разным количеством желтка в яйцеклетках: при его увеличении дробление становится сначала неравномерным, а затем (у рыб, птиц и рептилий) неполным поверхностным. В результате меняется и строение бластулы — имеется у видов с маленьким количеством желтка, — со средним и дискобластула — с большим. Кроме того, ход развития на ранних стадиях резко изменяется у наземных позвоночных в связи с появлением зародышевых оболочек .
Несмотря на то, что биогенетический закон не был подтвержден научными фактами, его широкая известность обусловлена тем, что он воспринимался как подтверждение дарвиновской теории эволюции . Однако он вовсе не следует из классического эволюционного учения.
Например, если вид А3 возник путём эволюции из более древнего вида А1 через ряд переходных форм (A1 => А2 => A3) , то, в соответствии с биогенетическим законом (в его модифицированном варианте), возможен и обратный процесс, при котором вид А3 превращается в А2 путём укорочения развития и выпадения его конечных стадий ( неотения или педогенез ).
Дарвинизм и синтетическая теория эволюции отрицают возможность полного возврата к предковым формам ( Закон необратимости эволюции Долло ). Причиной этого, в частности, являются перестройки эмбрионального развития на его ранних стадиях ( по А. Н. Северцову ), при которых генетические программы развития меняются настолько существенно, что их полное восстановление в ходе дальнейшей эволюции становится практически невероятным. Однако в случае неотенической эволюции вида не происходит возврата к предковой форме как таковой, поскольку зародыш определенного биологического вида отражает её лишь приблизительно. Отпадание конечных стадий развития при неотении не отменяет общего возрастания энтропии внутри системы. Таким образом полная картина такого примера эволюции следующая: A1 => А2 => A3 => А4, где А4 в чём-то схож с А2, но не равен ему. Например, гусеницы, способные к размножению без перехода ко взрослой стадии, подобны предкам насекомых, но имеют множество морфологических черт, существенно их отличающих.
Накопление фактов и теоретические разработки показали, что биогенетический закон в формулировке Геккеля в чистом виде никогда не выполняется. Рекапитуляция может быть только частичной. В XX веке эти факты заставили эмбриологов отвергнуть биогенетический закон в Геккелевских формулировках.
В каком-то смысле в биогенетическом законе перепутаны причины и следствия. Филогенез есть последовательность онтогенезов, следовательно, изменения взрослых форм в ходе филогенеза могут основываться только на изменениях онтогенеза. К такому пониманию соотношения онтогенеза и филогенеза биологи начали приходить еще в начале XX века.
В частности, А. Н. Северцов в 1912—1939 годах разработал теорию филэмбриогенезов . Согласно ей, все эмбриональные и личиночные признаки делятся на ценогенезы и филэмбриогенезы . Термин « ценогенез », предложенный Геккелем, Северцов трактовал иначе. Для Геккеля ценогенез (любые новые признаки, искажавшие рекапитуляцию) был противоположностью палингенеза (сохранения в развитии неизменных признаков, имевшихся и у предков). Северцов термином «ценогенез» обозначал признаки, которые служат приспособлениями к эмбриональному или личиночному образу жизни и у взрослых форм не встречаются, так как не могут иметь для них адаптивного значения. К ценогенезам Северцов относил, например, зародышевые оболочки амниот ( амнион , хорион , аллантоис ), плаценту млекопитающих, яйцевой зуб зародышей птиц и рептилий и др.
Филэмбриогенезы — это такие изменения онтогенеза, которые в ходе эволюции приводят к изменению признаков взрослых особей. Северцов разделил филэмбриогенезы на анаболии, девиации и архаллаксисы. Анаболия — удлинение онтогенеза, сопровождающееся надставкой стадий. Только при этом способе эволюции наблюдается рекапитуляция — признаки зародышей или личинок потомков напоминают признаки взрослых предков. При девиации происходят изменения на средних стадиях развития, что приводят к более резким изменениям в строении взрослого организма, чем при анаболии. При этом способе эволюции онтогенеза рекапитулировать признаки предковых форм могут лишь ранние стадии потомков. При архаллаксисах изменения происходят на самых ранних стадиях онтогенеза, изменения в строении взрослого организма наиболее часто существенны, а рекапитуляции невозможны.
В конце XX века опровержения биогенетического закона стали все более резкими. Например, С. Гилберт пишет: «Такая точка зрения ( о повторении онтогенезом филогенеза ) была научно дискредитирована даже раньше, чем была предложена… Она распространилась в биологии и общественных науках… прежде, чем было показано, что в её основе лежат ложные предпосылки» .
Столь же резко высказываются Р. Рэфф и Т. Кофмен : «Последний удар биогенетическому закону был нанесен тогда, когда стало ясно, что …морфологические адаптации имеют важное значение… для всех стадий онтогенеза» (с.31); «Вторичное открытие и развитие Менделевской генетики на рубеже двух столетий покажет, что в сущности биогенетический закон — это всего лишь иллюзия» (с.30),
Американский палеонтолог Нил Шубин отозвался о биогенетическом законе Геккеля следующим образом :
Новые данные, полученные за последние сто лет, подтверждают правоту скорее Бэра, чем Геккеля. Когда Геккель сравнивал эмбрионы одного вида с взрослыми особями другого, он во многом сравнивал круглое с красным. Развитие большинства животных отчасти действительно повторяет ход их эволюции, но для того, чтобы выявить механизмы эволюционных преобразований, плодотворнее сравнивать эмбрионы одного вида с эмбрионами другого, а не эмбрионы одного с взрослыми особями другого.
Оригинальный текст (англ.)In the past one hundred years, time and new evidence have treated von Baer much more kindly. In comparing embryos of one species to adults of another, Haeckel was comparing apples to oranges. A more meaningful comparison is one where we can ultimately uncover the mechanisms that drive evolution. For that, we compare embryos of one species to embryos of another.
|
|
|
|---|