Ядро Linux
- 1 year ago
- 0
- 0

Ядро одиночного пути ( лат. nucleus tractus solitarii ) — одно из ядер продолговатого мозга у человека и млекопитающих . Представляет собой тяж серого вещества . Отростки нейронов ядра входят в состав лицевого , языко-глоточного и блуждающего нервов . Ядро является местом входа чувствительных нервов от внутренних органов, служит переключателем вагусных рефлексов . Участвует в автономной регуляции сердечно-сосудистой , иммунной , пищеварительной и дыхательной систем . Благодаря мощным взаимосвязям с гипоталамусом является звеном в формировании аппетита .
Ядро одиночного пути находится в дорсо-медиальной части продолговатого мозга. Разные специалисты выделяют до 10 отделов ядра.
В ядре присутствуют нейроны различного размера и формы. Значительная часть нейронов — вставочные .
В отличие от других ядер, тела нейронов ядра одиночного пути содержат большинство известных нейромедиаторов млекопитающих.
Нейроны ядра имеют достаточно высокое количество рецепторов к грелину . Некоторые нейроны каудальной части экспрессируют белок проопиомеланокортин , а также имеют рецепторы к лептину и холецистокинину .
Афферентные пути идут в составе тройничного , языко-глоточного, лицевого и блуждающего нервов. К ядру идут окончания от барорецепторов каротидного синуса и аортальных телец. Также к ядру подходят и спинномозговые пути от грудных и поясничных сегментов. На него проецируются окончания от шовного ядра и ретикулярной формации , а также от вентральной зоны продолговатого мозга.
Главные нисходящие пути из переднего мозга начинаются в гипоталамических ядрах и в миндалине .
Эфферентные пути в основном обеспечивают обратную связь между ядром и органами, которые посылают к нему свои нервные окончания. Эти пути заканчиваются на:
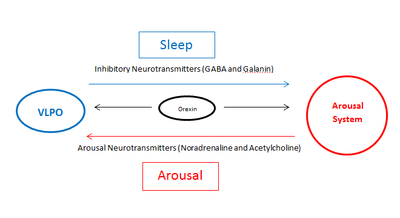
Ядро одинокого пути является центром барорецепции, вкуса, регуляции дыхания и желудочно-кишечной моторики. Также оно регулирует поступление жидкости из крови в спинномозговую жидкость .
Вкусовые рецепторы соединены синапсами с чувствительными нейронами узлов лицевого, языко-глоточного и блуждающего нерва. Аксоны этих нейронов достигают нервных клеток передней (ростральной) части ядра одиночного пути, которые иннервируют подкорковые центры вкуса в вентральных ядрах таламуса. При этом отростки с правого ядра переходят на левую часть таламуса и наоборот. Ядра таламуса передают вкусовую информацию далее к коре головного мозга.
Ядро регулирует сокращение гладких мышц желудка. Сенсорные нейроны узловатого ганглия реагируют на механический или химический раздражитель, который действует на стенку желудка, и выделяют возбуждающий медиатор глутамат , активируя нейроны ядра одиночного пути. Эти нейроны могут выделять возбуждающие (глутамат или норадреналин ) или тормозные ( ГАМК ) нейромедиаторы, воздействуя на мотонейроны заднего ядра блуждающего нерва. Заднее ядро аксонами своих нервных клеток возбуждает нейроны стенки желудка с помощью выделения ацетилхолина . Последние нейроны могут также секретировать ацетилхолин, возбуждая мышцы желудка, или выделять тормозные медиаторы: вазоинтестинальный пептид и оксид азота .
Другим механизмом в котором задействованы нейроны ядра, является формирование мигрирующего моторного комплекса желудка и кишки. Его началом является возбуждение окончаний вагуса серотонином , который выделяется энтерохромафинными клетками в полость желудка под влиянием гормона мотилина. Нервные импульсы достигают возбуждающих глутаматергических клеток ядра одинокого пути, которые возбуждают мотонейроны заднего ядра блуждающего нерва. Эти нейроны активируют нервные клетки стенки желудка, которые в ответ вызывают сокращение мышц желудка вследствие выделения ацетилхолина. Выделение серотонина стимулирует дальнейшую секрецию мотилина, поэтому процесс повторяется .
Нейроны ядра является нервным центром рефлекса Геринга-Бройера . Афферентные окончания от рецепторов растяжения лёгких производят нейромедиаторы глутамат и АТФ, которые возбуждают тормозные нейроны ядра, которые в свою очередь тормозят мотонейроны дыхательных мышц .
Ядро одиночного пути является составляющей дыхательного центра. Переднебоковая часть этого ядра состоит из инспираторных нейронов . От них направляется экстрапирамидный путь, связанный с регуляцией содружественности дыхательных мышц — одиночно-спинномозговой путь ( лат. tractus solitariospinalis ) .
Показано, что через ядро одиночного пути передается обратная связь при обучении пению . Нервные импульсы поступают от рецепторов гортани через блуждающий и язико-глоточный нервы, а затем передаются от ядра к вентральному заднемедиальному ядру таламуса . Оттуда информация достигает соматосенсорной коры. Также существует взаимная связь ядра одиночного пути с центральным серым веществом , где происходит консолидация сигналов, приводящих к созданию звуков .
Ядро также может участвовать в нервном контроле иммунной системы, а именно в так называемом «воспалительном рефлексе». Чувствительные окончания блуждающего нерва активируются провоспалительными цитокинами TNF-α и IL-1β и медиаторами гистамином и простагландинами . В ответ возбуждающие нейроны ядра одинокого пути активируют вагусные эффекторные окончания, которые выделяют ацетилхолин . Ацетилхолин связывается с альфа- никотиновыми рецепторами на макрофагах и блокирует производство провоспалительных цитокинов .
Исследования крыс со спонтанной гипертензией показало, что ядро одиночного пути в них продуцирует отличающиеся от обычных крыс количества медиаторов воспаления (простагландинов, лейкотриенов, интерлейкинов). Эти медиаторы побуждают лейкоциты двигаться к окружающим кровеносным сосудам, прикрепляться к эндотелию и вызвать воспаление мозга. Также у крыс с наследственной гипертензией показано уменьшение активности фермента лейкотриен-4В-12 гидроксидегидрогеназы, который разрушает эту молекулу, и увеличение активности фермента 5-липооксигеназы, которая синтезирует этот фактор из арахидоновой кислоты. Таким образом воспаление сосудов вблизи ядра одинокого пути может быть одной из причин артериальной гипертензии .
Нарушение работы ядра одиночного пути наблюдают при некоторых видах аутизма . Ядро является одним из немногих участков мозга, где гематоэнцефалический барьер является неполным в течение первого года жизни ребёнка после рождения, и в нём до того времени существуют фенестри, которые позволяют токсинам накапливаться в клетках ядра. Возможно, проявление аутизма после первого года жизни может быть связано именно с накоплением токсинов в ядре одиночного пути .
Также нейроны ядра могут быть задействованы во вторичных нарушениях моторики желудка вызванных повреждением спинного мозга .
Долгое время после открытия одиночного пути, ядра вокруг него не были исследованы. Анатомы считали, что тяж серого вещества вокруг одиночного пути состоит из нескольких отдельных ядер. Первое указание на ядро одинокого пути датируется 1895 годом (под названием «дорзальне сенсорное ядро языко-глоточного нерва»), а в 1897 году ядро впервые появляется в монографии под современным названием . Однако ещё много лет после этого многие анатомов считали, что блуждающий и языко-глоточный нерв начинаются где-то ещё. Окончательно топографию ядер вокруг одиночного пути установили лишь в 1960-е годы .
Долгое время в учебниках по анатомии и физиологии главным центром вкуса считалось вставочное ядро продолговатого мозга, поэтому нервные окончания лицевого нерва рисовали проходящими сквозь ядро одинокого пути. Лишь в 1980-е годы была выявлена роль вставного ядра в вестибулярных рефлексах, а центром вкуса было признано ядро одиночного пути .