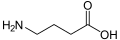
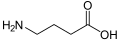
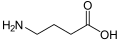
-Baclofen_Enantiomers_Structural_Formulae.png)

ГАМК-оперон
- 1 year ago
- 0
- 0


ГАМК B -рецептор (GABABR) — метаботропный трансмембранный G-белок-связанный подтип ГАМК-рецепторов , воздействующий через G-белки на калиевые ионные каналы клетки. Рецептор был открыт в 1981 году благодаря детальному фармакологическому анализу эффектов ГАМК в мозгу. Встречается во всех отделах вегетативной нервной системы и центральной нервной системы . Существует версия, что опьянение объясняется активацией этанолом ГАМК B2 рецептора.

Структурно ГАМК B -рецептор входит в одно суперсемейство с метаботропными глутаматными рецепторами . Две субъединицы, R1 и , судя по всему, образуют гетеродимеры , сцепляясь внутриклеточными С-концами . Первичная структура субъединиц схожа на 35 %. Каждая из субъединиц ГАМК B -рецептора имеет большой внеклеточный N-конец, за которым последовательно расположены семь трансмембранных доменов и внутриклеточный С-конец, который отвечает за гетеродимеризацию. Рецептор становится полностью функциональным только после гетеродимеризации. Более того, если указанные субъединицы экспрессируются раздельно, R1 неспособна достичь внешней клеточной мембраны, оставаясь на эндоплазматическом ретикулуме, но R2 неспособна связывать ГАМК. Несмотря на открытый полностью функциональный С-домен одиночной субъединицы R2, до сих пор не было получено свидетельств о возможности её сочетания со свободными субъединицами других аналогичных GPCR-рецепторов.
Всем представителям данного класса GPCR-рецепторов присущ сходный механизм активации, который известен под названием «Модель венериной мухоловки ». Построенные функциональные модели этой структуры предполагают, что лиганд попадает к N-концевому участку сайта связывания на субъединице R1, проходя между двух больших лопастей, способных к вращению благодаря специальному «шарнирному» участку (см. рисунок)
После того как агонист попадает в щель между подвижными частями субъединиц R1, эти части смыкаются и блокируют его. Аналогичная блокирующая структура «венериной мухоловки» присутствует и в R2-субъединице, но при этом у данной субъединицы отсутствуют аминокислотные остатки, необходимые для формирования места связывания агониста и/или антагониста. Возможно, субъединица R2 может быть активированной ещё не идентифицированным агонистом (неизвестно, эндогенным или экзогенным) — но несмотря на все опыты, такое активирующее вещество ( агонист ) до сих пор не найдено.
Благодаря активации ГАМК B -рецептора осуществляется регулирование нескольких комплексных процессов внутри нервной системы: сюда входит ингибирование аденилатциклазы; редукция агонист-стимулированного синтеза инозитол-1,4,5-трифосфат; ингибирование потенциал-зависимых Са 2+ -каналов, а также, по последним данным, активация К + -каналов. Передача эффекта с ГАМК B -рецепторов на аденилатциклазы осуществляется комплексом G-белков, а именно субъединицами Gαi и Gαo. Агонист-зависимое G-белковое связывание обеспечивается благодаря взаимодействию с одним (или более) внутриклеточным доменом субъединицы R2, как обычно полагают, второй и третьей внутриклеточными петлями (но все данные в пользу этого, пока, есть лишь косвенные). Калиевые каналы, действие которых регулируется ГАМК B -рецепторами, испытывают в данном случае воздействия через активацию βγ-комплекса G-белков (Gβγ). Этот же самый G-белковый комплекс функционально объединяет ГАМК B -рецепторы с пресинаптическими кальциевыми каналами N-, Р- и Т-типа. Это препятствует выделению нейромедиатора в синаптическую щель . Таким образом ГАМК B -рецептор является рецептором, тормозящим нейрон.
С помощью иммуногистохимических методов достоверно установлено, что как субъединица R1, так и субъединица R2 коэкспрессируются (то есть синтезируются совместно) практически во всех регионах мозга, хотя в распределении функциональных субъединиц наблюдаются некоторые различия. Например, субъединицы типа R1а преобладают в гипоталамусе , зрительной коре, стволе головного мозга и в гранулярном слое коры мозжечка . В то же время субъединицы типа R1b могут быть найдены в приповерхностных слоях коры больших полушарий, в спинном мозге и в молекулярном слое коры мозжечка.
Субъединица R2 найдена практически везде: в гиппокампе, коре больших полушарий, в клетках Пуркинье мозжечка и в спинном мозге — в соответствии с тем, что ГАМК B -рецепторы везде представлены гетеродимерами. Тем не менее, в некоторых пробах в опыте экспрессировалась только одна субъединица рецептора при отсутствии R2. R1-субъединица рецептора не может формировать функциональный рецептор; но, возможно, другие, ещё не идентифицированы субъединицы ГАМК B -рецептора могут связываться с ней при отсутствии R2 и формировать функционирующую рецепторную структуру.
Локализация ГАМК B -рецепторов в отдельном нейроне преимущественно внесинаптическая (за пределами синаптических структур). Это является признаком выполнения ингибиторной роли этими рецепторами только при очень интенсивном высвобождении ГАМК, когда она диффундирует за пределы синаптических щелей; также это хорошо согласуется с назначением ГАМК B -рецепторов, как одного из механизмов долговременной медленной модуляции синаптического ингибирования.

Материал из украинской и английской Википедии