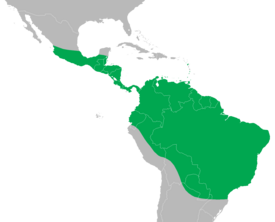
Наиболее известен благодаря наличию трёх разных типов самцов соревнующихся друг с другом по принципу
камень, ножницы, бумага
— каждый тип имеет преимущества над следующим по кругу.
Содержание
Название
Название
stansburiana
дано в честь
, который собрал первые образцы во время экспедиции 1849—1851 годов для изучения и исследования
Большого Соленого озера
в
штате Юта
.
Систематика
Самец с сине-жёлтой окраской и характерным тёмным пятном за предплечьем
В систематике этих ящериц есть много спорного.
Бесчисленное количество форм были описаны как подвиды и даже виды.
Формы, которые встречаются в большей части Мексики (за исключением
Нижней Калифорнии
) признаны другим видом
Uta stejnegeri
.
Ящерицы с
(англ.)
(
и
острова Седрос
ранее были выделены в отдельные виды
Uta stellata
и
Uta concinna
, но теперь входят в
Uta stansburiana
.
Ящерицы с островов Санта-Каталина и Салсипуедес, так называемые
Uta squamata
и
Uta antiqua
иногда также включаются в этот вид. Это точно не верно в последнем случае, и скорее всего неверно в первом.
Популяцию с Южной
Нижней Калифорнии
, вероятно, следует также выделить в вид
Uta elegans
.
Предложенные подвиды
martinensis
и
taylori
, вероятно, излишни.
Популяция с
Анхель-де-ла-Гуарда
, Мехия и Раса острова почти наверняка представляют собой отдельный вид ближе к
Uta palmeri
.
Популяция с острова Сан-Эстебана скорее всего ближе к
squamata
.
Популяции с Лас-Энкантадас названы
Uta encantadae
,
Uta lowei
и
Uta tumidarostra
но статус не решён окончательно; эти отдельные популяции имеют относительно позднее происхождение и иногда включаются в этот вид, но их уникальными приспособлениями к жизни предполагают, что они должны считаться разными; как один или как три вида остается нерешенным.
Описание
Самцы могут вырастать до 60 мм, самки обычно немного меньше. Степень пигментации зависит от пола и популяции.
Некоторые самцы имеют голубые пятна на спинах и хвостах, а их бока могут быть жёлтыми или оранжевыми, другие не имеют окраски. Самки могут иметь полосы вдоль спины и боков, или почти не иметь окраски.
Оба пола имеют заметное пятно на боку, сразу за передней конечностью.
Окраска тесно связана с брачным поведением самцов и самок.
Полиморфизм
Самцы
Самцы имеют ярко выраженный
полиморфизм
по окраске горла, и могут быть разделены на три типа:
оранжевогорлые
,
синегорлые
и
желтогорлые
.
Число особей каждого из этих трёх типов варьирует. Равновесие поддерживается механизмом
камень-ножницы-бумага
— наименьший по численности тип самцов в определенном сезоне спаривается чаще и имеет наибольшее количество потомства в следующем году.
Оранжевогорлые самцы ультрадоминантны. Они самые крупные и агрессивные. Контролируют большие территории, около 100 м
2
. Держат гаремы самок. Они способны отнять самку у синегорлых самцов, но уязвимы для желтогорлых.
Оранжевогорлые самцы также значительно чаще погибают по сравнению с двумя другими типами.
Синегорлые самцы являются доминантными. Они являются промежуточными по размеру, и контролируют небольшую территорию, с единственной самкой. Они в состоянии защитить её от желтогорлых, но не в силах противостоять натиску оранжевогорлых самцов.
Желтогорлые самцы имеют окраску схожую с окраской зрелых самок. При столкновении с доминирующим оранжевогорлым или синегорлым самцом, они как правило имитируют поведение самки. В отличие от других типов, желтогорлые самцы не контролируют территории. Они обитают в широких диапазонах перекрывающимся с территориями нескольких других самцов.
Они вступают в вязки с чужими самками на чужих территориях. Этого легче добиться в гареме оранжевогорлого самца, чем у единственной самки синегорлого. Хотя оранжевогорлые самцы имеют высокий уровень смертности, желтогорлые самцы имеют более высокие относительные показатели посмертных родов.
Желтогорлые самцы могут в некоторых случаях превращаться в синегорлых в течение сезона размножения. Это преобразование, как правило, вызвано смертью доминантного самца по соседству. При этом голубые вкрапления желтогорлых самцов качественно отличаются от синих пятен генетически синегорлых. Не все желтогорлые самцы трансформируются, но если это случается, они отказываются от поведенческого подражания самкам и принимают доминантный тип поведения. Превращений в другом направлении, в которых доминирующие самцы становятся желтогорлыми зафиксировано не было.
Самки
Поведение самок также коррелирует с окраской горла. Оранжевогорлые самки используют
R-стратегию
. Они, как правило, производят большие кладки, состоящие из множества маленьких яиц.
В отличие от них, жёлтогорлые самки используют
K-стратегию
— откладывают меньше, более крупных яиц.
Как и у самцов, численность этих двух типов самок циклична во времени. Однако, цикл короче — два года по сравнению с 4—5 годами у самцов.
Оранжевогорлые самки более успешны при низкой плотности населения, где конкуренция за пищу менее жестока и давление от
хищничества
не столь значительно.
Когда плотность населения высока, и достаточно хищников, жёлтогорлые самки, как правило, имеют более высокий репродуктивный успех — их более крупные потомки имеют более высокие краткосрочную и долгосрочную выживаемость, и эти преимущества усиливаются в периоды дефицита.
Генетика
Тип самцов определяется генетически, и имеет высокую
наследуемость
.
Он определяется одним
Менделевским признаком
с тремя аллелями. У самцов,
o
-аллель
доминантный
, и
b
-аллель является рецессивный к
y
-аллелю.
Фенотипически
оранжевогорлый самец имеет
генотип
либо
oo
,
ob
, или
oy
. Желтогорлые самцы имеют генотипы либо
yy
или
yb
, и синегорлый самец исключительно
bb
.
Самки, с доминирующим
o
-аллелем оранжевогорлые,
а у тех, что без
о
-аллеля развиваться жёлтое горло.
Вязка
Поведение
Самки откладывают в среднем по 5,1 яйца, максимум 9 яиц, в одной кладке. Меньшие кладки часто ассоциируется с жёлтогорлыми самками.
Агрессии и ухаживания
Доминантные самцы проявляют агрессивность в обороне своих территорий.
При появлении чужаков они приходят в состояние повышенной бдительности, выполняют одно или несколько отжиманий — вертикальных покачиваний, выгибают спину, и раздвигают конечности при приближении непрошеного гостя.
Если нарушитель другой самец, владелец территории устремляется к нарушителю, который затем, как правило убегает.
Если это самка, то владелец инициирует ухаживания, состоящие из обходов, покусываний, лизания, обнюхивания, подпрыгиваний, и в итоге
совокупления
. Форма тела и пассивность являются основным
релизером
для ухаживания. Самцы были замечены в попытках совокупляться с небольшими ящерицами других видов.
Длина хвоста важна в определении
доминирования иерархии
.
Как и у многих других ящериц, пятнистые ящерицы используют
хвостовую автототомию
в качестве эвакуационного механизма. Однако, уменьшение длины хвоста влечёт потерю социального статуса для самцов и для самок.
Самцы используют автотомию менее охотно, чем самки, вероятно, из-за возросшей значимости социального статуса для самцов — репродуктивный успех напрямую связан с их социальным статусом.
Питание
Питание зависит от
пола
или
сезона
. В ходе исследования, проведенного Бэст и другими их диета в значительной степени основана на
членистоногих
.
Исследования показывают корреляцию между полом и диетой. Половые различия в пищевом поведении может действовать в пользу снижения внутривидовой конкуренции за ресурсы.
↑
Sinervo, B.; C.M. Lively.
The rock–paper–scissors game and the evolution of alternative male strategies
(англ.)
// Nature : journal. — 1996. —
Vol. 380
,
no. 6571
. —
P. 240—243
. —
doi
:
.
↑
Sinervo, Barry; Paul Licht.
Proximate Constraints on the Evolution of Egg Size, Number, and Total Clutch Mass in Lizards
(англ.)
// Science : journal. — 1991. —
Vol. 252
,
no. 5010
. —
P. 1300—1302
. —
doi
:
.
Moll, Edward. 2005.
Grismer, L. L.
(англ.)
// Herpetologica : journal. — 1994. —
Vol. 50
. —
P. 451—474
.
Upton, Darlene E.; Murphy, Robert W.
Phylogeny of the side-blotched lizards (Phrynosomatidae:
Uta
) based on mtDNA sequences: support for midpeninsular seaway in Baja California
(англ.)
//
Molecular Phylogenetics and Evolution
: journal. —
Academic Press
, 1997. —
Vol. 8
,
no. 1
. —
P. 104—113
. —
doi
:
.
Schmidt, Karl Patterson.
(англ.)
// American Museum Novitates : journal. — 1921. —
Vol. 22
. —
P. 1—6
.
14 июня 2007 года.
Collins, Joseph T.
(англ.)
// Herpetological Review : journal. — 1991. —
Vol. 22
,
no. 2
. —
P. 42—43
.
29 сентября 2007 года.
Murphy, Robert W. & Aguirre-León, Gustavo (2002): The Nonavian Reptiles: Origins and Evolution.
↑
Tinkle, D.W.
(англ.)
// University of Michigan Museum of Zoology: Miscellaneous Publications : journal. — 1967. —
No. 132
.
↑
Alonzo, S.H.; Barry Sinervo.
Mate choice games, context-dependent good genes, and genetic cycles in the side-blotched lizard, Uta stansburiana
(англ.)
// Behavioral Ecology and Sociobiology : journal. — 2001. —
Vol. 49
,
no. 2—3
. —
P. 176—186
. —
doi
:
.
↑
Sinervo, Barry; Donald B. Miles; W.Anthony Frankino; Matthew Klukowski; Dale F. DeNardo.
Testosterone, Endurance, and Darwinian Fitness: Natural and Sexual Selection on the Physiological Bases of Alternative Male Behaviors in Side-Blotched Lizards
(англ.)
// Hormones and Behavior : journal. — 2000. —
Vol. 38
,
no. 4
. —
P. 222—233
. —
doi
:
. —
.
Ferguson, Gary W.; Stanley F. Fox.
(англ.)
//
Evolution
: journal. —
Wiley-VCH
, 1984. —
Vol. 38
,
no. 2
. —
P. 342—349
. —
doi
:
.
Ferguson, Gary W.
Releasers of courtship and territorial behaviour in the side blotched lizard Uta stansburiana
(англ.)
// Animal Behavior : journal. — 1966. —
Vol. 14
,
no. 1
. —
P. 89—92
. —
doi
:
.
Fox, Stanley F.; Nancy A. Heger; Linda S. Delay.
Social cost of tail loss in Uta stansburiana: lizard tails as status-signalling badges
(англ.)
//
(англ.)
(
: journal. —
Elsevier
, 1990. —
Vol. 39
,
no. 3
. —
P. 549—554
. —
doi
:
.
Fox, Stanley F.; Jason M. Conder; Allie E. Smith.
Sexual Dimorphism in the Ease of Tail Autotomy: Uta stansburiana with and without Previous Tail Loss
(англ.)
//
(англ.)
(
: journal. —
(англ.)
(
, 1998. —
Vol. 1998
,
no. 2
. —
P. 376—382
. —
doi
:
.
Best, Troy L.; A. L. Gennaro.
Feeding Ecology of the Lizard, Uta stansburiana, in Southeastern New Mexico
(англ.)
// Journal of Herpetology : journal. — 1984. — September (
vol. 18
,
no. 3
). —
P. 291—301
. —
doi
:
.
Ссылки
: Cooperation between unrelated male lizards adds a new wrinkle to evolutionary theory (Essay on Sinervo & Clobert 2001 paper)