Interested Article - Аллели

- 2021-03-01
- 1
Алле́ли (от греч. ἀλλήλων — друг друга, взаимно; сокр. от аллеломорфы ) — различные формы одного и того же гена , расположенные в одинаковых участках ( локусах ) гомологичных хромосом , определяют направление развития конкретного признака . В диплоидном организме может быть два одинаковых аллеля одного гена, в этом случае организм называется гомозиготным , или два разных, что приводит к гетерозиготному организму.
Нормальные диплоидные соматические клетки содержат два аллеля одного гена (по числу гомологичных хромосом), а гаплоидные гаметы — лишь по одному аллелю каждого гена. Для признаков, подчиняющихся законам Менделя , можно рассматривать доминантные и рецессивные аллели. Если генотип особи содержит два разных аллеля (особь — гетерозигота ), проявление признака зависит только от одного из них — доминантного. Рецессивный же аллель влияет на фенотип , только если находится в обеих хромосомах (особь — гомозигота ). Таким образом, доминантный аллель подавляет рецессивный. В более сложных случаях наблюдаются другие типы аллельных взаимодействий (см. ниже) .
Однако, несмотря на разнообразие взаимодействия аллелей, порой весьма сложных, все они подчиняются первому закону Менделя — закону единообразия гибридов первого поколения .
Этимология
Обычно считают, что слово «аллель» впервые было предложено Вильгельмом Иогенсеном в 1909 году в монографии « Elementen der exakten Erblichkeitslehre », однако на самом деле в этом издании употреблялся термин «аллеломорф» . Термин аллель Иогенсен впервые использовал на английском в статье в журнале « Hereditas » в 1923 году , а на немецком в 1926 году — в третьем издании своей монографии . Слово происходит от др.-греч. ἀλλήλων , что означает «друг друга». На русском языке одним из первых слово «аллель» использовал Константин Андреевич Фляксбергер в 1935 году в переводе книги Грегора Менделя. Большинство слов русского языка заимствованных из немецкого языка, оканчивающиеся на ударное « -el », относятся к женскому роду. Однако анализ употребления этого слова в современной научной литературе показывает на устойчивую тенденцию употребления этого слова в мужском роде .
Обозначение аллелей
Обычно в качестве обозначения аллеля применяют сокращение названия соответствующего гена до одной или нескольких букв. В случае наличия только 2 аллелей, чтобы отличить доминантный аллель от рецессивного, первую букву в обозначении доминантного пишут заглавной.
Если же имеет место , то используется следующая система обозначения аллелей, например, в случае нижеописанной мутации ade у дрожжей. Ответственный за неё ген обозначают заглавными латинскими буквами: ADE 2 . Множественные аллели этого гена записывают так: ade 2—1 , ade 2—2 и т. д. Сам же признак, как это принято в генетике микроорганизмов , обозначают ADE .
При этом само слово «аллель» употребляется в русском языке как в женском , так и в мужском роде .
Типы аллельных взаимодействий
Вскоре после переоткрытия законов Менделя было установлено, что взаимодействие генов не исчерпывается одним лишь полным доминированием доминантного аллеля над рецессивным. В действительности между аллелями одного гена и различными генами в генотипе существуют сложные и разнообразные взаимодействия, играющие немаловажную роль в реализации информации , заключённой в каждом отдельном гене .
Доминантность

Доминирование (доминантность) заключается в том, что один из аллелей пары (доминантный) маскирует или полностью подавляет проявление второго аллеля (рецессивного). При этом степень подавления рецессивного признака обусловливает различные варианты доминирования:
- Полное доминирование — взаимодействие двух аллелей одного гена, когда доминантный аллель полностью исключает проявление действия второго аллеля. В фенотипе присутствует только признак, задаваемый доминантным аллелем. Например, в экспериментах Менделя пурпурная окраска цветка полностью доминировала над белой .
- Неполное доминирование — доминантный аллель в гетерозиготном состоянии не полностью подавляет действие рецессивного аллеля. Гетерозиготы имеют промежуточный характер признака. Например, если в гомозиготном состоянии один аллель определяет красную окраску цветка, а другой — белую, то гетерозиготный гибрид будет иметь розовые цветки . В некоторых источниках неполное доминирование характеризуют как такой тип взаимодействия аллелей, когда признак у гибридов F 1 занимает не среднее положение, а отклоняется в сторону родителя с доминирующим признаком. Полностью же средний вариант (как, например, приведённый выше пример наследования окраски цветков) относят к промежуточному характеру наследования , то есть отсутствию доминирования .
- Сверхдоминирование — более сильное проявление признака у гетерозиготной особи, чем у любой гомозиготной . На этом типе аллельного взаимодействия основано явление гетерозиса (превосходство над родителями по жизнеспособности, энергии роста, плодовитости, продуктивности) .
- Кодоминирование — проявление у гибридов нового варианта признака, обусловленного взаимодействием двух разных аллелей одного гена. При этом, в отличие от неполного доминирования, оба аллеля проявляются в полной мере. Наиболее известным примером является наследование групп крови у человека . Некоторые источники также понимают именно под кодоминированием отсутствие доминантно-рецессивных отношений .
- Доминирование, связанное с полом , происходит тогда, когда один и тот же аллель у самцов проявляется как доминантный, а у самок — как рецессивный. Например, у овцематок доминирует комолость (R), а у баранов — рогатость (R 1 ) .
Другие типы аллельных взаимодействий
Межаллельная комплементация

Межаллельная комплементация — довольно редкое взаимодействие аллелей . Она имеет место в том случае, когда возможно формирование нормального признака D у организма, гетерозиготного по двум мутантным аллелям D 1 D 2 .
Установлено, что все гены, аллели которых взаимодействуют таким образом, контролируют структуру ферментов , построенных из одинаковых белковых субъединиц. Если же мутантные аллели отличаются тем, что кодируют полипептиды различных доменов , то при их объединении в гетерозиготе ( компаунде ) в молекуле фермента соберутся все необходимые функциональные центры (так как мутантные аллели ответственны за дефектность лишь некоторых из них), то ферментативная активность, а значит, и нормальный фенотип будут восстановлены .
Примером данного типа взаимодействия аллелей является проявление мутации ade у дрожжей , заключающейся в отсутствии активности фермента . При этом мутанты имеют красный цвет, обусловленный тем, что из-за отсутствия этого фермента они вынуждены поглощать экзогенный аденин и субстрат вместе с ним, а последний, накапливаясь в клетках , придаёт им красную окраску. У нормальных форм сохраняется белый цвет .
Аллельное исключение
Аллельное исключение происходит при инактивации одной из Х-хромосом у особей гомогаметного пола , приводящего в соответствие дозы Х-генов у всех представителей пола (к примеру, у женщин в каждой клетке экспрессируется лишь одна Х-хромосома, другая же переходит в неактивный гетерохроматин (т. н. тельце Барра ). Таким образом, женский организм у человека является мозаичным, так как в разных клетках могут экспрессироваться разные хромосомы). При этом проявляются только аллели, находящиеся в активной Х-хромосоме, те же аллели, которые находятся в инактивированной, как бы исключаются и не проявляются в фенотипе . При этом в каждой клетке экспрессируется или один, или другой аллель, в зависимости от того, какая Х-хромосома была инактивирована.
Схожая картина наблюдается в В-лимфоцитах , синтезирующих антитела к определённым антигенам . Поскольку эти иммуноглобулины моноспецифичны, каждый лимфоцит должен выбрать между экспрессией отцовского или материнского аллеля .
Множественный аллелизм
Множественным аллелизмом называют явление существование в популяции более двух аллелей данного гена. Одним из наиболее известных проявлений множественного аллелизма является уже упоминавшееся наследование групп крови у человека: у человека имеется ген I , попарные комбинации трёх различных аллелей которого — I A , I B , i — приводят к развитию той или иной группы крови у человека. Взаимодействие множественных аллелей может быть различным. Так, в вышеописанном примере аллели I A и I B доминируют над аллелью i , а при одновременном наличии в генотипе аллелей I A и I B они взаимодействуют друг с другом по типу кодоминирования .
Множественный аллелизм широко распространён в природе. Так, по этому механизму определяется тип совместимости при опылении у высших растений , типы спаривания у грибов , окраска шерсти у животных, цвет глаз у дрозофилы, форма рисунка на листьях белого клевера . Кроме того, у растений, животных и микроорганизмов имеются — белковые молекулы, различия между которыми определяются аллелями одного гена .
Летальные аллели

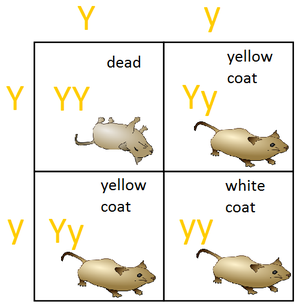
Летальными называются аллели, носители которых погибают из-за нарушений развития или заболеваний, связанных с работой данного гена. Эти аллели возникают тогда, когда нормальная работа гена нарушается из-за мутации , произошедшей в некотором его аллеле. Летальное действие таких аллелей может проявляться как в эмбриональном , так и постэмбриональном периоде жизни организма. Примером летального аллеля, из-за которой организм погибает на стадии эмбриона , является летальный аллель Y-гена, участвующего в определении окраски шерсти у мышей (см. рисунок) . В качестве летального аллеля, чьё действие проявляется во взрослом возрасте, может быть рассмотрен доминантный аллель, ответственный за развитие хореи Гентингтона у человека .
Частоты аллелей и генотипов
В популяции бесконечно большого размера, в которой не действует естественный отбор , не идёт мутационный процесс, отсутствует обмен особями с другими популяциями, не происходит дрейф генов , все скрещивания случайны — частоты генотипов по какому-либо гену (в случае, если в популяции есть два аллеля этого гена) будут поддерживаться постоянными из поколения в поколение и соответствовать уравнению

где — доля гомозигот по одному из аллелей, — частота этого аллеля, — доля гомозигот по альтернативному аллелю, — частота соответствующего аллеля, — доля гетерозигот .





См. также
Примечания
- Гл. Ред. А. М. Прохоров. Большая Советская Энциклопедия. — 3. — М. : Советская Энциклопедия, 1970. — Т. 1. — С. 445. — 608 с.
- (англ.) . www.etymonline.com . Дата обращения: 2 октября 2023. 28 марта 2023 года.
- , с. 43—55.
- ↑ , с. 45—55.
- ↑ Синюшин А. А. К истории генетической терминологии: какого рода слово «аллель»? // Экологическая генетика : журнал. — 2016. — Т. 14 , № 1 . — С. 49—54 . — doi : .
- Johannsen W. Some remarks about units in heredity (англ.) // Hereditas : journal. — 1923. — Vol. 4 , no. 1—2 . — P. 133—141 . — doi : .
- . slovari.ru (17 февраля 2004). Дата обращения: 27 августа 2012. 29 февраля 2012 года.
- ↑ Максимов Г.В., Василенко В.Н., Кононенко О.И., Максимов А.Г., Максимов В.Г. Сборник задач по генетике. — М. : «Вузовская книга», 2010. — С. 15—20. — 144 с. — 300 экз. — ISBN 978-5-9502-0420-3 .
- ↑ Ярыгин В. Н., Васильева В. И., Волков И. Н., Синельщикова В. В. Биология. В 2 кн / Ярыгин В. Н.. — М. : Высшая школа, 2010. — Т. 1. — С. 157—158. — 432 с.
- Билич Г.Л., Крыжановский В.А. Биология. Полный курс: В 4 т. — издание 5-е, дополненное и переработанное. — М. : Издательство Оникс, 2009. — Т. 1. — С. 726. — 864 с. — ISBN 978-5-488-02311-6 .
- . Дата обращения: 8 июня 2014. Архивировано из 16 июня 2014 года.
- Neil A. Campbell, Jane B. Reece, Lisa A. Urry, Michael L. Cain et. al. Biology (англ.) . — 9th edition. — Benjamin Cummings. — P. 278. — 1263 p. — ISBN 978-0-321-55823-7 .
- , с. 551.
Литература
- Инге-Вечтомов С. Г. Генетика с основами селекции. — СПб. : Издательство Н-Л, 2010. — 718 с. — ISBN 978-5-94869-105-3 .
Ссылки
| Ключевые понятия |

|
|
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
|
|
Эта статья входит в число
добротных статей
русскоязычного раздела Википедии.
|
- 2021-03-01
- 1