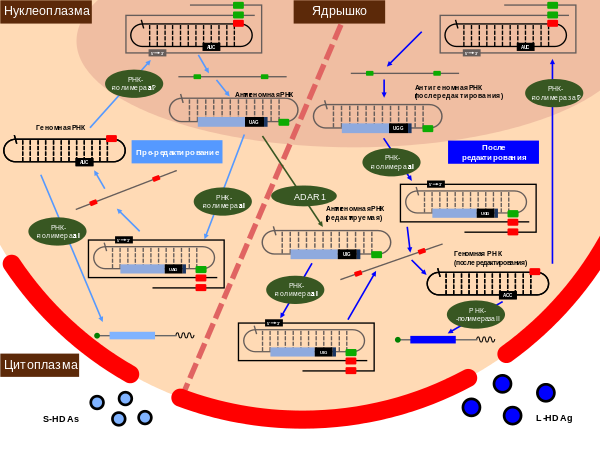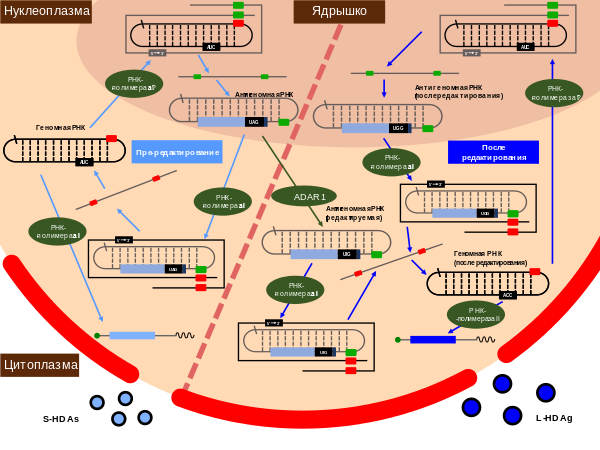
Вирус гепатита дельта
- 1 year ago
- 0
- 0

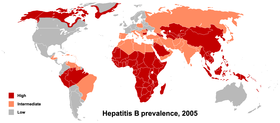
Ви́рус гепати́та B (вирус гепатита b, «б»; англ. Hepatitis B virus , HBV) — ДНК-содержащий вирус из семейства гепаднавирусов , возбудитель вирусного гепатита B . В мире по различным оценкам от 3 до 6 % людей инфицировано вирусом гепатита B. Носительство вируса не обязательно сопровождается гепатитом, однако носитель вируса может заражать других людей, а также животных.
Несмотря на то, что существует вакцина для предотвращения гепатита В, вирус гепатита В остаётся глобальной проблемой здравоохранения. Гепатит В может протекать остро, а затем переходить в хроническую форму, приводя к другим заболеваниям и состояниям здоровья . В дополнение к возникновению гепатита, заражение вирусом гепатита В может привести к циррозу и гепатоцеллюлярной карциноме .
Также было высказано предположение, что заболевание может увеличить риск возникновения рака поджелудочной железы .

Вирус гепатита В классифицируется в роде , который содержит 11 других видов. Род классифицируется как часть семейства Hepadnaviridae , в которое входят четыре других рода: , , and . Это семейство вирусов является единственным представителем вирусного отряда Blubervirales .
Вирусы, сходные с гепатитом В, были обнаружены у всех человекообразных обезьян ( орангутанов , гиббонов , горилл и шимпанзе ), у обезьян Старого Света , и у шерстистых обезьян Нового Света (вирус гепатита В шерстистых обезьян), что позволяет предположить древнее происхождение этого вируса у приматов.
Вирус подразделяется на четыре основных серотипа (adr, adw, ayr, ayw) на основе антигенных эпитопов , присутствующих на белках его оболочки . Эти серотипы основаны на общей детерминанте (a) и двух взаимоисключающих парах детерминант (d/y и w/r). Вирусные штаммы также были разделены на десять генотипов (A-J) и сорок подгенотипов в соответствии с общей вариацией нуклеотидной последовательности генома . Генотипы имеют чёткое географическое распределение и используются для отслеживания эволюции и передачи вируса. Различия между генотипами влияют на тяжесть заболевания, течение и вероятность осложнений, и ответ на лечение . Серотипы и генотипы не обязательно соответствуют.
У летучих мышей выделен ряд пока ещё не классифицированных видов, подобных гепатиту В .
Вирус гепатита В является членом семейства гепаднавирусов . Вирусная частица, называемая частицей Дейна ( вирион ), состоит из внешней липидной оболочки и икосаэдрического ядра нуклеокапсида , состоящего из белка . Нуклеокапсид содержит вирусную ДНК и ДНК-полимеразу , которая обладает обратной транскриптазной активностью, подобной ретровирусам . Внешняя оболочка содержит встроенные белки, которые участвуют в связывании вируса с восприимчивыми клетками и проникновении в них. Вирус является одним из самых маленьких оболочечных вирусов животных с диаметром вириона 42 нм, но также существуют формы, в том числе нитевидные и сферические тела, лишённые ядра. Эти частицы не являются инфекционными и состоят из липида и белка, которые образуют часть поверхности вириона, который называется поверхностным антигеном ( ) и вырабатывается в избытке в течение жизненного цикла вируса .
Вирус состоит из:
Вирусу гепатита D требуются частицы оболочки HBV, чтобы стать вирулентными .
Раннюю эволюцию вируса гепатита B, как и всех других вирусов, трудно установить. Выявление гепаднавирусов у широкого круга позвоночных предполагает длительную коэволюцию. Идентификация эндогенных элементов hepadnaviridae, общих для различных видов птиц, показывает присутствие этого вируса у птиц в течение по меньшей мере 70 миллионов лет . Хотя аналогичные доказательства отсутствуют для млекопитающих, филогенетическое положение ортогепаднавирусов как сестринской клады авигепаднавирусов предполагает присутствие вируса в предоках амниот и последующая коэволюция как с птицами, так и с млекопитающими после их расхождения (> 300 миллионов лет назад). Также было высказано предположение, что Новый мировой гепаднавирус летучих мышей может быть источником гепаднавирусов приматов . У авихепаднавирусов отсутствует Х-белок, но в геноме утиного гепаднавируса присутствует рудиментарная Х-рамка считывания . Х-белок, возможно, произошёл от ДНК-гликозилазы.
Недавно реконструкция геномов вируса гепатита В по древним человеческим останкам позволила более детально исследовать эволюцию этого вируса у людей . В 2021 году исследование реконструировало 137 древних геномов вируса гепатита В и доказало присутствие вируса у людей, по крайней мере, с 10 000 лет . Самый последний общий предок всех известных человеческих линий вируса гепатита В датируется от 20 000 до 12 000 лет назад. Однако нельзя сказать, присутствовал ли вирус у людей задолго до этого или был приобретён незадолго до этого от другого вида животных. Было показано, что эволюция вируса гепатита В у людей отражает известные события человеческой истории, такие как первое заселение Америки в позднем плейстоцене и переходный период неолита в Европе . Эти исследования также показали, что некоторые древние штаммы вируса гепатита В все ещё заражают людей, в то время как другие вымерли . Штаммы ВГВ, обнаруженные у африканских и юго-восточноазиатских обезьян (шимпанзе, горилл, орангутанов и гиббонов), по-видимому, родственны человеческим штаммам ВГВ, что может отражать прошлые случаи межвидовой передачи .
Исследование изолятов из циркумполярной арктической человеческой популяции показало, что предок субгенотипа B5 (эндемичного типа, обнаруженного в этой популяции), что предковый вирус возник в Азии около 2000 лет назад (95 % HPD 900 г. до н. э. — 830 г. н. э.) . Слияние произошло около 1000 г. н. э. Этот субгенотип распространился из Азии первоначально в Гренландию , а затем распространился на запад в течение последних 400 лет.
Геном вируса гепатита В состоит из кольцевой ДНК , но это необычно, потому что ДНК не является полностью двухцепочечной. Один конец полноразмерной цепи связан с вирусной ДНК-полимеразой . Длина генома составляет 3020-3320 нуклеотидов (для цепи полной длины) и 1700—2800 нуклеотидов (для короткой цепи) .
Отрицательная (некодирующая) цепь комплементарна вирусной мРНК. Вирусная ДНК обнаруживается в ядре вскоре после инфицирования клетки. Частично двухцепочечная ДНК становится полностью двухцепочечной путём завершения (+) смысловой цепи клеточными ДНК-полимеразами (вирусная ДНК−полимераза используется на более поздней стадии) и удаления белка вирусной полимеразы (P) из (-) смысловой цепи и короткой последовательности РНК из(+) смысловой нити. Некодирующие основания удаляются из концов (−) смысловой цепи, и концы соединяются.
Вирусные гены транскрибируются клеточной РНК-полимеразой II в ядре клетки с ковалентно замкнутой кольцевой ДНК (cccDNA). В геноме HBV были идентифицированы два усилителя, обозначенные как enhancer I (EnhI) и enhancer II (EnhII). Оба усилителя проявляют большую активность в клетках печёночного происхождения, и вместе они управляют и регулируют экспрессию полных вирусных транскриптов .
Существует четыре известных гена, кодируемых геномом, которые называются C, P, S и X. Основной белок кодируется геном C (HBcAg), и его стартовому кодону предшествует стартовый кодон AUG, из которого вырабатывается прекоординатный белок. HBeAg образуется в результате протеолитической переработки белка, предшествующего ядру. ДНК-полимераза кодируется геном P. Ген S — это ген, который кодирует поверхностный антиген (HBsAg). Ген HBsAg представляет собой одну длинную открытую рамку считывания, но содержит три «стартовых» (ATG) кодона в рамке, которые делят ген на три секции: pre-S1, pre-S2 и S. Из-за множества стартовых кодонов полипептиды трёх разных размеров называются большими, средними и малыми (pre-S2).-Вырабатываются S1 + pre-S2+ S, pre-S2+ S или S) .
Функция белка, кодируемого геном X, до конца не изучена , но некоторые данные свидетельствуют о том, что он может функционировать как транскрипционный трансактиватор. Интересно, что слитый белок с Х-ядром 40 кДа кодируется длинным вирусным транскриптом размером 3,9 кб, функция которого остаётся неясной . Синтез 3.9 кб РНК инициируется в промоторной области гена X, и транскрипт полиаденилируется только после второго раунда транскрипции. Аналогичное поведение характерно и для других видов длинных прегеномных/предъядерных (pg/pc) РНК. Таким образом, механизм вирусной транскрипции должен игнорировать сигнал poly (A) на первом раунде транскрипции.
В геноме HBV было идентифицировано несколько некодирующих элементов РНК . К ним относятся: , и сигнал инкапсидации РНК .
Известно восемь генотипов, обозначенных от А до H . Был описан возможный новый генотип «I» , но принятие этого обозначения не является универсальным .
Различия между генотипами составляют более 8 %. Генотипы A и D распространены повсеместно; генотипы C и B характерны для Юго-Восточной Азии и Японии. Генотип Е распространён преимущественно в Африке. Генотип F был найден среди коренного населения Южной Америки и на Аляске. Генотип G спорадически встречается в различных частях света, в частности в Соединённых Штатах Америки и во Франции. Генотипы Е и G характеризуются низкой вариабельностью в последовательности нуклеотидов в геноме, по сравнению с другими генотипами.
Генотипы вируса гепатита B могут обладать различными биологическими свойствами. В последнее время все большое значение придаётся генотипу вируса в клинических аспектах течения вирусной инфекции, а также чувствительности к антивирусным препаратам. На сегодняшний день установлено, что инфекция, вызванная вирусом гепатита B генотипов В и С, коррелирует с повреждением печени; а инфекция, обусловленная вирусом гепатита B генотипа А, эффективно излечивается терапевтическими методами с использованием интерферона.
Генотип D имеет 10 подгенотипов .

Жизненный цикл вируса гепатита В сложен. Гепатит В является одним из немногих известных неретровирусных вирусов, которые используют обратную транскрипцию как часть процесса репликации.
HBV обладает способностью .
Упомянутый выше поверхностный HBsAg-антиген, находящийся в наружной оболочке, применяют для производства высокоэффективной профилактической вакцины .
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)