Липопротеины
- 1 year ago
- 0
- 0


Липопротеи́ны ( липопротеиды ) — класс сложных белков , простетическая группа которых представлена каким-либо липидом . Так, в составе липопротеинов могут быть свободные жирные кислоты , нейтральные жиры, фосфолипиды , холестериды.
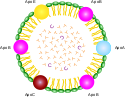
Липопротеины представляют собой комплексы, состоящие из белков ( аполипопротеинов ; сокращенно — апо-ЛП) и липидов, связь между которыми осуществляется посредством гидрофобных и электростатических взаимодействий.
Липопротеины подразделяют на свободные, или растворимые в воде (липопротеины плазмы крови , молока и др.), и нерастворимые, т. е. структурные (липопротеины мембран клетки, миелиновой оболочки нервных волокон, хлоропластов растений).
Среди свободных липопротеинов (они занимают ключевое положение в транспорте и метаболизме липидов) наиболее изучены липопротеины плазмы крови, которые классифицируют по их плотности. Чем выше содержание в них липидов, тем ниже плотность липопротеинов. Различают липопротеины высокой плотности (ЛВП), низкой плотности (ЛНП), очень низкой плотности (ЛОНП) и хиломикроны. Каждая группа липопротеинов очень неоднородна по размерам частиц (наиболее крупные — хиломикроны ) и содержанию в ней аполипопротеинов. Все группы липопротеинов плазмы содержат полярные и неполярные липиды в разных соотношениях.
| Вид | Размеры | Функция |
|---|---|---|
| Липопротеины высокой плотности (ЛВП) | 8-11 нм | Транспорт холестерина от периферийных тканей к печени |
| Липопротеины низкой плотности (ЛНП) | 18-26 нм | Транспорт холестерина, триацилглицеридов и фосфолипидов от печени к периферийным тканям |
| Липопротеины промежуточной (средней) плотности ЛПП (ЛСП) | 25-35 нм | Транспорт холестерина, триацилглицеридов и фосфолипидов от печени к периферийным тканям |
| Липопротеины очень низкой плотности (ЛОНП) | 30-80 нм | Транспорт холестерина, триацилглицеридов и фосфолипидов от печени к периферийным тканям |
| Хиломикроны | 75-1200 нм | Транспорт холестерина и жирных кислот, поступающих с пищей, из кишечника в периферические ткани и печень |
Нековалентная связь в липопротеинах между белками и липидами имеет важное биологическое значение. Она обусловливает возможность свободного обмена липидов и модуляцию свойств липопротеинов в организме.
Липопротеины являются:

Хиломикроны образуются в лимфатической системе ворсинок кишечника. Они переносят до половины всех триацилглицеролов и холестерина лимфы. Новосинтезированные хиломикроны содержат интегральный белок В-48. Апопротеин В встраивается в липопротеины в гЭПР, где синтезируются триацилглицеролы. В аппарате Гольджи к белкам добавляются углеводы. Они высвобождаются из клеток кишечника обратным пиноцитозом . После этого хиломикроны поступают в лимфатические сосуды ворсинок и уносятся лимфой. Попадая в кровоток, они получают апопротеины С и Е от ЛВП. На стенках капилляров находится липопротеинлипаза (ЛПЛ) (прикрепляется к ним протеогликановыми цепями гепарансульфата). В печени также есть своя липаза , но она менее эффективно атакует хиломикроны. Апопротеин С2 активирует липопротеинлипазу, которая расщепляет триглицериды хиломикрона до ди- и моноглицеридов, а затем — до свободной жирной кислоты и глицерола. Жирные кислоты транспортируются в мышечные и жировые ткани или связываются с альбумином в крови. По мере липолиза хиломикроны теряют большинство своих триацилглицеролов, относительное содержание холестерина и его эфиров увеличивается. Диаметр остатка хиломикрона уменьшается. Апопротеин С2 возвращается на ЛВП, апопротеин Е сохраняется. Остатки хиломикронов поглощаются печенью. Поглощение осуществляется через рецепторный эндоцитоз , с помощью рецепторов апопротеина Е. В печени эфиры холестерина и триацилглицеролы окончательно гидролизуются.
ЛОНП переносят триацилглицеролы, а также фосфолипиды, холестерин и его эфиры из печени в другие ткани. Метаболизм ЛОНП похож на метаболизм хиломикронов. Интегральным белком их является другой апопротеин В, В-100. ЛОНП высвобождаются из клеток печени обратным пиноцитозом, после чего через слой эпителиальных клеток поступают в капилляры печени. В крови на них переносятся апопротеины С2 и Е с ЛВП. Триацилглицеролы ЛОНП, как в случае с хиломикронами, расщепляются при активации ЛПЛ с помощью апопротеина С2, свободные жирные кислоты поступают в ткани. По мере расщепления триацилглицеролов диаметр ЛОНП уменьшается, и они превращаются в ЛПП. Эфир-холестерин-переносящий белок (апопротеин D в составе ЛВП) переносит на ЛОНП эфиры холестерина от ЛВП в обмен на фосфолипиды и триглицериды .
Половина ЛПП поглощается печенью с помощью рецепторного эндоцитоза через рецепторы апопротеина Е и B-100. Триацилглицериды ЛПП гидролизуются печёночной липазой. Апопротеины С2 и Е возвращаются на ЛВП. частица превращается в ЛНП. Относительное содержание холестерина в ЛНП значительно увеличивается, диаметр частицы сокращается. (Они также переносят триглицериды, каротиноиды, витамин Е и др.) ЛНП поглощаются клетками печени (70%) и внепечёночных тканей с помощью рецепторного эндоцитоза. Однако лигандом теперь, в основном, служит белок В-100. Рецептор называется «рецептором ЛНП».
ЛВП обеспечивают обратный транспорт холестерина из внепечёночных тканей к печени. ЛВП синтезируются в печени. В новообразованных ЛВП содержатся апопротеины А1 и А2. Апопротеин А1 синтезируется также в кишечнике, где входит в состав хиломикронов, но при липолизе в крови быстро переносятся на ЛВП. Апопротеин С синтезируется в печени, выделяется в кровоток и уже в кровотоке переносится на ЛВП. Новообразованный ЛВП похож на диск: фосфолипидный бислой, включающий свободный холестерин и апопротеин. Апопротеин А1 — активатор фермента лецитинхолестеринацилтрансферазы (ЛХАТ). Этот фермент связан с поверхностью ЛВП в плазме крови. ЛХАТ катализирует реакцию между фосфолипидом ЛВП и свободным холестерином частицы. При этом образуются эфиры холестерина и лизолецитин. Неполярные эфиры холестерина перемещаются внутрь частицы, освобождая место на поверхности для захвата нового холестерина, лизолецитин — на альбумин крови. Неполярное ядро раздвигает бислой , ЛВП приобретает сферическую форму. Этерифицированный холестерин переносится с ЛВП на ЛОНП, ЛНП и хиломикроны специальным белком ЛВП — переносчиком эфиров холестерола (апопротеин D), в обмен на фосфолипиды и триглицериды. ЛВП поглощается клетками печени с помощью рецепторного эндоцитоза через рецептор апопротеина Е.
Специфичности рецепторов апопротеинов Е и В-100 частично пересекаются. Они находятся на поверхности мембран клеток в клатриновой кавеоле. При соединении с лигандами кавеола замыкается в везикулу и липопротеин эндоцитируется. В лизосомах эфиры холестерина гидролизуются и холестерин поступает в клетку.
|
Липиды
:
|
|
|---|---|
|
Структуры
клеточной мембраны
|
|
|---|---|
| Мембранные белки | |
| Другое | |