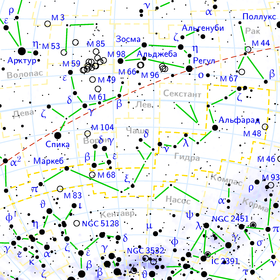
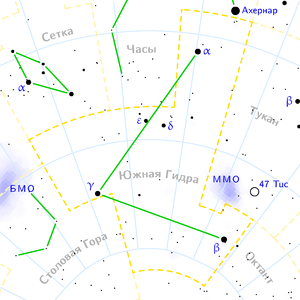
Гамма Гидры
- 1 year ago
- 0
- 0

Гидры ( лат. Hydra ) — род пресноводных сидячих кишечнополостных из класса гидроидных (Hydrozoa). Представители обитают в стоячих водоёмах и реках с медленным течением, прикрепляясь к водным растениям или грунту. Длина тела гидры составляет 1—20 мм, иногда несколько более. Это одиночный малоподвижный полип .
Гидры (пресноводные полипы) впервые были описаны швейцарским натуралистом XVIII века Абраамом Трамбле . Открытие вначале вызвало недоверие биологов, особенно способность полипов к регенерации . В 1744 году Трамбле выпустил двухтомную монографию, посвящённую полипам .
Тело гидры цилиндрической формы, на переднем конце тела (на околоротовом конусе, или гипостоме) расположен рот, окружённый венчиком из 5—12 щупалец. У некоторых видов тело разделено на туловище и стебелёк. На заднем конце тела (стебелька) расположена подошва, с её помощью гидра передвигается и прикрепляется к чему-либо. Гидра обладает радиальной (одноосно-гетеропольной) симметрией. Ось симметрии соединяет два полюса — оральный, на котором находится рот, и аборальный, на котором находится подошва. Через ось симметрии можно провести несколько плоскостей симметрии, разделяющих тело на две зеркально симметричных половины.
Тело гидры — мешок со стенкой из двух слоёв клеток ( эктодермы и энтодермы ), между которыми находится тонкий слой межклеточного вещества ( мезоглея ). Полость тела гидры — гастральная полость — образует выросты, заходящие внутрь щупалец . Хотя обычно считают, что у гидры есть только одно ведущее в гастральную полость отверстие (ротовое), на самом деле на подошве гидры имеется узкая аборальная пора. Через неё может выделяться жидкость из кишечной полости, а также пузырёк газа. При этом гидра вместе с пузырьком открепляется от субстрата и всплывает, удерживаясь вниз головой в толще воды. Таким способом она может расселяться по водоёму. Что касается ротового отверстия, то у непитающейся гидры оно фактически отсутствует — клетки эктодермы ротового конуса смыкаются и образуют плотные контакты, такие же, как и на других участках тела . Поэтому при питании гидре каждый раз приходится «прорывать» рот заново.

Эпителиально-мускульные клетки эктодермы и энтодермы образуют основную массу тела гидры. У гидры около 20 000 эпителиально-мускульных клеток.
Клетки эктодермы имеют цилиндрическую форму эпителиальных частей и формируют однослойный покровный эпителий . К мезоглее прилегают сократимые отростки данных клеток, образующие продольную мускулатуру гидры.
Эпителиально-мускульные клетки энтодермы направлены эпителиальными частями в полость кишки и несут по 2—5 жгутиков , которые перемешивают пищу. Эти клетки могут образовывать ложноножки, с помощью которых захватывают частицы пищи. В клетках формируются пищеварительные вакуоли.
Эпителиально-мускульные клетки эктодермы и энтодермы представляют собой две независимые клеточные линии. В верхней трети туловища гидры они делятся митотически, а их потомки постепенно смещаются либо в сторону гипостома и щупалец, либо в сторону подошвы. По мере перемещения происходит дифференцировка клеток: так, клетки эктодермы на щупальцах дают клетки стрекательных батарей, а на подошве — железистые клетки, выделяющие слизь.
Железистые клетки энтодермы выделяют в гастральную полость пищеварительные ферменты, которые расщепляют пищу. Эти клетки образуются из интерстициальных клеток. У гидры около 5000 железистых клеток.
Между эпителиально-мускульными клетками находятся группы мелких, округлых клеток, называемых промежуточными, или интерстициальными (i-клетки). У гидры их около 15 000. Это недифференцированные клетки. Они могут превращаться в остальные типы клеток тела гидры, кроме эпителиально-мускульных. Промежуточные клетки обладают всеми свойствами мультипотентных стволовых клеток. Доказано, что каждая промежуточная клетка потенциально способна дать как половые, так и соматические клетки. Стволовые промежуточные клетки не мигрируют, однако их дифференцирующиеся клетки-потомки способны к быстрым миграциям.
Нервные клетки образуют в эктодерме примитивную диффузную нервную систему — рассеянное нервное сплетение (диффузный плексус). В энтодерме есть отдельные нервные клетки. Всего у гидры около 5000 нейронов . У гидры имеются сгущения диффузного плексуса на подошве, вокруг рта и на щупальцах. По новым данным, у гидры (по крайней мере у некоторых видов) имеется околоротовое нервное кольцо, сходное с нервным кольцом, расположенным на крае зонтика у гидромедуз.
У гидры нет четкого деления на чувствительные, вставочные и моторные нейроны. Одна и та же клетка может воспринимать раздражение и передавать сигнал эпителиально-мускульным клеткам. Тем не менее есть два основных типа нервных клеток — чувствительные и ганглиозные. Тела чувствительных клеток расположены поперек эпителиального пласта, они имеют неподвижный жгутик, окружённый воротничком из микроворсинок, который торчит во внешнюю среду и способен воспринимать раздражение. Ганглиозные клетки расположены в основании эпителиально-мускульных, их отростки не выходят во внешнюю среду. По морфологии большинство нейронов гидры — биполярные или мультиполярные.
В нервной системе гидры присутствуют как электрические, так и химические синапсы . Из нейромедиаторов у гидры обнаружены дофамин, серотонин, норадреналин, гамма-аминомасляная кислота, глютамат, глицин и многие нейропептиды (вазопрессин, вещество Р и др.).
Гидра — наиболее примитивное животное, в нервных клетках которого обнаружены чувствительные к свету белки опсины . Анализ гена опсина гидры позволяет предположить, что опсины гидры и человека имеют общее происхождение .
Стрекательные клетки образуются из промежуточных только в области туловища. Сначала промежуточная клетка делится 3-5 раз, образуя кластер (гнездо) из предшественников стрекательных клеток (книдобластов), соединённых цитоплазматическими мостиками. Затем начинается дифференцировка, в ходе которой мостики исчезают. Дифференцирующиеся книдоциты мигрируют в щупальца. Стрекательные клетки наиболее многочисленные из всех клеточных типов, их у гидры около 55 000.
Стрекательная клетка имеет стрекательную капсулу, заполненную ядовитым веществом. Внутрь капсулы ввёрнута стрекательная нить. На поверхности клетки находится чувствительный волосок, при его раздражении нить выбрасывается и поражает жертву. После выстреливания нити клетки погибают, а из промежуточных клеток образуются новые.
У гидры есть четыре типа стрекательных клеток — стенотелы (пенетранты), десмонемы (вольвенты), голотрихи изоризы (большие глютинанты) и атрихи изоризы (малые глютинанты). При охоте первыми выстреливают вольвенты. Их спиральные стрекательные нити опутывают выросты тела жертвы и обеспечивают её удержание. Под действием рывков жертвы и вызванной ими вибрации срабатывают имеющие более высокий порог раздражения пенетранты. Шипы, имеющиеся у основания их стрекательных нитей, заякориваются в теле добычи, а через полую стрекательную нить в её тело вводится яд.
Большое количество стрекательных клеток находится на щупальцах, где они образуют стрекательные батареи. Обычно в состав батареи входит одна крупная эпителиально-мускульная клетка, в которую погружены стрекательные клетки. В центре батареи находится крупная пенетранта, вокруг неё — более мелкие вольвенты и глютинанты. Книдоциты соединены десмосомами с мускульными волокнами эпителиально-мускульной клетки. Большие глютинанты (их стрекательная нить имеет шипы, но не имеет, как и у вольвент, отверстия на вершине), видимо, в основном используются для защиты. Малые глютинанты используются только при передвижении гидры для прочного прикрепления щупальцами к субстрату. Их выстреливание блокируется экстрактами из тканей жертв гидры.
Выстреливание пенетрант гидры было изучено с помощью сверхвысокоскоростной киносъёмки. Оказалось, что весь процесс выстреливания занимает около 3 мс. В его начальной фазе (до выворачивания шипов) скорость его достигает 2 м/c, а ускорение составляет около 40 000 g (данные 1984 года ); видимо, это один из самых быстрых клеточных процессов, известных в природе. Первым видимым изменением (менее чем через 10 мкс после стимуляции) было увеличение объёма стрекательной капсулы примерно на 10 %, затем объём снижается почти до 50 % от исходного. В дальнейшем выяснилось, что и скорость, и ускорение при выстреливании нематоцист были сильно недооценены; по данным 2006 года , на ранней фазе выстреливания (выбрасывание шипов) скорость этого процесса составляет 9-18 м/с, а ускорение составляет от 1 000 000 до 5 400 000 g. Это позволяет нематоцисте массой около 1 нг развивать на кончиках шипов (диаметр которых составляет около 15 нм) давление порядка 7 гПа, что сравнимо с давлением пули на мишень и позволяет пробивать толстую кутикулу жертв.
Как и всем животным, гидрам свойственна оогамия . Большинство гидр раздельнополы, но встречаются гермафродитные линии гидр. И яйцеклетки, и сперматозоиды образуются из i-клеток. Считается, что это особые субпопуляции i-клеток, которые можно отличить по клеточным маркерам и которые в небольшом количестве присутствуют у гидр и в период бесполого размножения.
При оогенезе ооциты фагоцитируют целые оогонии, а затем несколько ооцитов сливаются, после чего ядро одного из них превращается в ядро яйцеклетки, а остальные ядра дегенерируют. Эти процессы обеспечивают быстрый рост яйцеклетки.
Как недавно показано, при сперматогенезе имеет место программированная клеточная смерть части клеток-предшественников сперматозоидов и их фагоцитирование окружающими клетками эктодермы
Дыхание и выделение продуктов обмена происходит через всю поверхность тела животного. Вероятно, в выделении некоторую роль играют вакуоли, которые есть в клетках гидры. Главная функция вакуолей, вероятно, осморегуляторная ; они выводят излишки воды, которые постоянно поступают в клетки гидры путём осмоса .
Гидры имеют сетчатую нервную систему. Наличие нервной системы позволяет гидре осуществлять простые рефлексы . Гидра реагирует на механическое раздражение, температуру, освещённость , наличие в воде химических веществ и на ряд других факторов внешней среды.
Гидра питается мелкими беспозвоночными —
дафниями
и другими ветвистоусыми,
циклопами
, а также олигохетами-наидидами. Есть данные о потреблении гидрами
коловраток
и
церкарий
трематод
. Добыча захватывается щупальцами с помощью стрекательных клеток, яд которых быстро парализует мелких жертв. Координированными движениями щупалец добыча подносится ко рту, а затем с помощью сокращений тела гидра «надевается» на жертву. Пищеварение начинается в кишечной полости (полостное пищеварение), заканчивается внутри пищеварительных вакуолей эпителиально-мускульных клеток энтодермы (внутриклеточное пищеварение). Непереваренные остатки пищи выбрасываются через рот.
Так как у гидры нет транспортной системы, а мезоглея (слой межклеточного вещества между экто- и энтодермой) достаточно плотная, возникает проблема транспорта питательных веществ к клеткам эктодермы. Эта проблема решается за счёт образования выростов клеток обоих слоёв, которые пересекают мезоглею и соединяются через
щелевые контакты
. Через них могут проходить мелкие органические молекулы (моносахариды, аминокислоты), что обеспечивает питание клеток эктодермы.
При благоприятных условиях гидра размножается бесполым путём. На теле животного (обычно в нижней трети туловища) образуется почка, она растет, затем формируются щупальца и прорывается рот. Молодая гидра отпочковывается от материнского организма (при этом материнский и дочерний полипы прикрепляются щупальцами к субстрату и тянут в разные стороны) и ведёт самостоятельный образ жизни. Осенью гидра переходит к половому размножению. На теле, в эктодерме закладываются гонады — половые железы, а в них из промежуточных клеток развиваются половые клетки. При образовании гонад гидр формируется медузоидный узелок. Это позволяет предполагать, что гонады гидры — сильно упрощённые споросаки, последний этап в ряду преобразования утраченного медузоидного поколения в орган. Большинство видов гидр раздельнополы, реже встречается гермафродитизм . Яйцеклетки гидр быстро растут, фагоцитируя окружающие клетки. Зрелые яйцеклетки достигают диаметра 0,5—1 мм. Оплодотворение происходит в теле гидры: через специальное отверстие в гонаде сперматозоид проникает к яйцеклетке и сливается с ней. Зигота претерпевает полное равномерное дробление , в результате которого образуется . Затем в результате смешанной деламинации (сочетание иммиграции и деламинации) осуществляется гаструляция . Вокруг зародыша формируется плотная защитная оболочка (эмбриотека) с выростами-шипиками. На стадии гаструлы зародыши впадают в анабиоз . Взрослые гидры погибают, а зародыши опускаются на дно и зимуют. Весной продолжается развитие, в паренхиме энтодермы путём расхождения клеток образуется кишечная полость, затем формируются зачатки щупалец, и из-под оболочки выходит молодая гидра. Таким образом, в отличие от большинства морских гидроидных, у гидры отсутствуют свободноплавающие личинки, развитие у неё прямое.
В норме у взрослой гидры клетки всех трёх клеточных линий интенсивно делятся в средней части тела и мигрируют к подошве, гипостому и кончикам щупалец. Там происходит гибель и слущивание клеток. Таким образом, все клетки тела гидры постоянно обновляются. При нормальном питании «избыток» делящихся клеток перемещается в почки, которые обычно образуются в нижней трети туловища.
Гидра обладает очень высокой способностью к регенерации . При разрезании поперек на несколько частей каждая часть восстанавливает «голову» и «ногу», сохраняя исходную полярность — рот и щупальца развиваются на той стороне, которая была ближе к оральному концу тела, а стебелек и подошва — на аборальной стороне фрагмента. Целый организм может восстанавливаться из отдельных небольших кусочков тела (менее 1/200 объёма), из кусочков щупалец, а также из взвеси клеток. При этом сам процесс регенерации не сопровождается усилением клеточных делений и представляет собой типичный пример морфаллаксиса .
Гидра может регенерировать из взвеси клеток, полученных путём мацерации (например, при протирании гидры через ). В экспериментах показано, что для восстановления головного конца достаточно образования агрегата из примерно 300 эпителиально-мускульных клеток. Показано, что регенерация нормального организма возможна из клеток одного слоя (только эктодермы или только энтодермы).
Фрагменты разрезанного тела гидры сохраняют информацию об ориентации оси тела организма в структуре актинового цитоскелета : при регенерации ось восстанавливается, волокна направляют деление клеток. Изменение структуры актинового скелета может привести к нарушениям в регенерации (образованию нескольких осей тела) .
Уже ранние опыты Абраама Трамбле показали, что при регенерации сохраняется полярность фрагмента. Если разрезать тело гидры поперек на несколько цилиндрических фрагментов, то на каждом из них ближе к бывшему оральному концу регенерируют гипостом и щупальца (в экспериментальной эмбриологии гидры закрепился термин «голова» для обозначения орального конца тела), а ближе к бывшему аборальному полюсу — подошва («нога»). При этом у тех фрагментов, которые располагались ближе к «голове», быстрее регенерирует «голова», а у располагавшихся ближе к «ноге» — «нога».
Позднее опыты по изучению регенерации были усовершенствованы в результате применения методики сращивания фрагментов разных особей. Если вырезать из боковой стороны туловища гидры фрагмент и срастить его с телом другой гидры, то возможны три исхода опыта: 1) фрагмент полностью сливается с телом реципиента; 2) фрагмент образует выступ, на конце которого развивается «голова» (то есть превращается в почку); 3) фрагмент образует выступ, на конце которого образуется «нога». Выяснилось, что процент образования «голов» тем выше, чем ближе к «голове» донора взят фрагмент для пересадки и чем дальше от «головы» реципиента он помещен. Эти и аналогичные опыты привели к постулированию существования четырёх веществ-морфогенов, регулирующих регенерацию — активатора и ингибитора «головы» и активатора и ингибитора «ноги». Эти вещества, согласно данной модели регенерации, образуют концентрационные градиенты: в районе «головы» у нормального полипа максимальна концентрация как активатора, так и ингибитора головы, а в районе «ноги» — максимальна концентрация и активатора, и ингибитора ноги.
Эти вещества действительно были обнаружены. Активатор головы — пептид из 11 аминокислот (pGlu-Pro-Pro-Gly-Gly-Ser-Lys-Val-Ile-Leu-Phe), активный в пикомолярной концентрации (первоначально 20 мкг активатора были получены из 200 кг морских актиний). У человека он присутствует в гипоталамусе и кишечнике и в той же концентрации обладает нейротрофическим действием. У гидры и млекопитающих этот пептид обладает также митогенным действием и влияет на клеточную дифференцировку.
Активатор ноги — тоже пептид с молекулярной массой, близкой к 1000 Да. Ингибиторы головы и ноги — низкомолекулярные гидрофильные вещества небелковой природы. В норме все четыре вещества выделяются нервными клетками гидры. Активатор головы имеет большее время полужизни (около 4 ч), чем ингибитор (30 мин) и медленнее диффундирует, так как связан с белком-носителем. Ингибитор головы в очень низкой концентрации подавляет выделение активатора, а в 20 раз большей концентрации — своё собственное выделение. Ингибитор ноги также ингибирует выделение активатора ноги.
При регенерации, как и при росте и бесполом размножении, эпителиально-мускульные клетки делятся самостоятельно, причем клетки эктодермы и энтодермы — две независимые клеточные линии. Остальные типы клеток (нервные, стрекательные и железистые) развиваются из промежуточных. Убив делящиеся промежуточные клетки высокой дозой радиации или колхицином , можно получить «безнервных», или эпителиальных гидр — они продолжают расти и почковаться, но отделяющиеся почки лишены нервных и стрекательных клеток. Культуру таких гидр удается поддерживать в лаборатории с помощью «насильственного» кормления.
Известны также мутантные линии «безнервных» гидр, у которых нет промежуточных клеток и у которых промежуточные клетки могут давать только сперматозоиды, но не соматические клетки, а также мутантные линии, у которых промежуточные клетки погибают при повышенной температуре.
«Безнервные» гидры сохраняют способность к регенерации.
Ещё в конце XIX века была выдвинута гипотеза о теоретическом бессмертии гидры, которую пытались научно доказать или опровергнуть на протяжении всего XX века . В 1997 году гипотеза была доказана экспериментальным путём Даниэлем Мартинесом . Эксперимент продолжался порядка четырёх лет и показал отсутствие смертности среди трёх групп гидр вследствие старения . Считается, что «бессмертность» гидр напрямую связана с их высокой регенерационной способностью.
Перед наступлением зимы, после перехода к половому размножению и созреванию покоящихся стадий, гидры в водоёмах средней полосы погибают. Видимо, это происходит не из-за нехватки пищи или непосредственного воздействия иных неблагоприятных факторов. Это говорит о наличии у гидр механизмов старения .
В водоёмах России и Украины наиболее часто встречаются следующие виды гидр (в настоящее время многие зоологи выделяют кроме рода Hydra ещё 2 рода — Pelmatohydra и Chlorohydra ):

У так называемых «зеленых» гидр Hydra (Chlorohydra) viridissima в клетках энтодермы живут эндосимбиотические водоросли рода Chlorella — . На свету такие гидры могут длительное время (более четырёх месяцев) обходиться без пищи, в то время как искусственно лишённые симбионтов гидры без кормления погибают через два месяца. Зоохлореллы проникают в яйцеклетки и передаются потомству трансовариально . Другие виды гидр в лабораторных условиях иногда удается заразить зоохлореллами, однако устойчивого симбиоза при этом не возникает.
Именно с наблюдений за зелёными гидрами начал свои исследования А. Трамбле .
На гидр могут нападать мальки рыб, для которых ожоги стрекательных клеток, видимо, довольно чувствительны: схватив гидру, малёк обычно выплёвывает её и отказывается от дальнейших попыток съесть.
На поверхности тела гидр в качестве паразитов или комменсалов часто обитают Kerona polyporum , триходина и другие инфузории .
К питанию тканями гидр приспособлен ветвистоусый рачок из семейства хидорид Anchistropus emarginatus .
Тканями гидр могут также питаться турбеллярии микростомы , которые способны использовать непереваренные молодые стрекательные клетки гидр в качестве защитных клеток — клептокнид .
Видимо, впервые описал гидру Антонио ван Левенгук . Подробно изучил питание, движение и бесполое размножение, а также регенерацию гидры Авраам Трамбле , который описал результаты своих опытов и наблюдений в книге «Мемуары к истории одного рода пресноводных полипов с руками в форме рогов» (первое издание вышло на французском языке в 1744 г.). Открытие Трамбле приобрело громкую славу, его опыты обсуждались в светских салонах и при французском королевском дворе. Эти опыты опровергли господствовавшее тогда убеждение, что отсутствие бесполого размножения и развитой регенерации у животных — одно из важнейших их отличий от растений. Считается, что изучение регенерации гидры (опыты А. Трамбле) положило начало экспериментальной зоологии . Научное название роду в соответствии с правилами зоологической номенклатуры присвоил Карл Линней . Название содержит отсылку к многоголовой Лернейской гидре , победа над которой была одним из двенадцати подвигов Геракла . Вероятно, Линней имел в виду регенерационные способности: когда Лернейской гидре отрубали одну голову, на её месте тут же вырастала другая .

В последние десятилетия гидра используется как модельный объект для изучения регенерации и процессов морфогенеза. Геном гидры (североамериканский вид Hydra magnipapillata ) частично расшифрован. В Японии и Германии есть коллекции мутантных линий гидры. Разработана методика получения трансгенных гидр.