Ритм-танец
- 1 year ago
- 0
- 0


Цирка́дные (циркадиа́нные) ри́тмы (от лат. circa «около, кругом» + dies «день») — циклические колебания интенсивности различных биологических процессов, связанные со сменой дня и ночи . Период циркадных ритмов обычно близок к 24 часам.
Несмотря на связь с внешними стимулами, циркадные ритмы имеют эндогенное происхождение, представляя, таким образом, биологические часы организма .
Циркадные ритмы присутствуют у таких организмов, как цианобактерии , грибы , растения , животные .
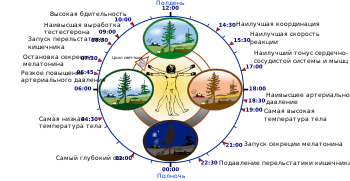
Самым известным циркадным ритмом является ритм сон - бодрствование .
Впервые об изменении положения листьев в течение дня у тамаринда ( Tamarindus indicus ) упоминает описывавший походы Александра Македонского .
В Новое время в 1729 году французский астроном Жан-Жак де Меран сообщил о ежедневных движениях листьев у мимозы стыдливой ( Mimosa pudica ). Эти движения повторялись с определённой периодичностью даже если растения помещались в темноту, где отсутствовали такие внешние стимулы как свет, что позволило предположить эндогенное происхождение биологических ритмов, к которым были приурочены движения листьев растения. Де Меран предположил, что эти ритмы могут иметь что-то общее с чередованием сна и бодрствования у человека.
Декандоль в 1834 году определил, что период, с которыми растения мимозы совершают данные листовые движения, короче длины суток и составляет примерно 22—23 часа.
В 1880 году Чарльз Дарвин и его сын Фрэнсис сделали предположение о наследственной природе циркадных ритмов. Предположение о наследственной природе циркадных ритмов было подтверждено окончательно опытами, во время которых скрещивались растения фасоли , периоды циркадных ритмов которых различались. У гибридов длина периода отличалась от длины периода у обоих родителей.
Эндогенная природа циркадных ритмов была окончательно подтверждена в 1984 году во время опытов с грибами вида Нейроспора густая ( Neurospora crassa ), проведёнными в космосе. Эти опыты показали независимость околосуточных ритмов от геофизических сигналов, связанных с вращением Земли вокруг своей оси.
В 1970-е годы Сеймур Бензер и его ученик Рональд Конопка изучали, можно ли идентифицировать гены, которые контролируют циркадный ритм у плодовых мух . Они продемонстрировали, что мутации неизвестного гена нарушают циркадные часы мух. Неизвестный ген получил название ген периода — Per (от англ. period ).
В 1984 году Джеффри Холл и Майкл Росбаш , работающие в тесном сотрудничестве в Брандейском университете в Бостоне , и Майкл Янг из Рокфеллеровского университета в Нью-Йорке смогли выделить ген Per . Затем Джеффри Холл и Майкл Росбаш обнаружили, что белок PER, кодируемый геном Per , накапливается в течение ночи и деградирует в течение дня. Таким образом, уровень белка PER колеблется в течение суток синхронно с циркадным ритмом. Учёные предположили, что белок PER блокирует активность гена Per . Они обосновали, что с помощью ингибирующей петли обратной связи белок может препятствовать своему собственному синтезу и тем самым регулировать собственный уровень в непрерывном циклическом ритме. Однако, чтобы блокировать активность гена Per , белок PER, который продуцируется в цитоплазме, должен был каким-то образом достигнуть клеточного ядра, где расположен генетический материал, — этот вопрос оставался невыясненным.
В 1994 году Майкл Янг обнаружил второй «часовой ген» циркадного ритма, timeless , кодирующий белок TIM, который требовался для нормального циркадного ритма. Майкл Янг показал, что когда белок TIM связан с белком PER, оба белка могут проникать в ядро клетки, где они блокируют активность гена Per , таким образом замыкая ингибирующую петлю обратной связи. Майкл Янг идентифицировал ещё один ген, doubletime , кодирующий белок DBT, который задерживал накопление белка PER. Совместное действие обнаруженных генов обеспечило понимание, как корректируется циркадный ритм для более точного соответствия 24-часовому циклу.
В последующие годы были выяснены другие молекулярные компоненты механизма, объясняющие его стабильность и функционирование. Были определены дополнительные белки, необходимые для активации гена Per , а также механизм, посредством которого свет может синхронизировать цикл.
В 2017 году Джеффри Холл, Майкл Росбаш и Майкл Янг были удостоены Нобелевской премии за открытие молекулярных механизмов, контролирующих циркадный ритм .
Циркадные ритмы растений связаны со сменой дня и ночи и важны для адаптации растений к суточным колебаниям таких параметров как температура, освещение, влажность. Растения существуют в постоянно меняющемся мире, поэтому циркадные ритмы важны для того, чтобы растение могло дать надлежащий ответ на абиотический стресс. Изменение положения листьев в течение суток — лишь один из многих ритмических процессов у растений. В течение суток колеблются такие параметры, как активность ферментов, интенсивность газообмена и фотосинтетическая активность.
В способности растений распознавать чередование дня и ночи играет роль фитохромная система. Примером работы такой системы является ритм цветения у растения Pharbitis nil . Цветение у этого растения зависит от длины светового дня: если день короче определённого интервала, то растение цветет, если длиннее — вегетирует. В течение суток условия освещения меняются из-за того, что солнце находится под разными углами к горизонту, и соответственно меняется спектральный состав света, что воспринимается различными фитохромами которые возбуждаются светом с разной длиной волны. Так, вечером в спектре много дальних красных лучей, которые активизируют только фитохром А, давая растению сигнал о приближении ночи. Получив этот сигнал, растение принимает соответствующие меры. Важность фитохромов для температурной адаптации была выяснена во время опытов с трансгенными осинами Populus tremula , у которых продукция фитохрома A была повышена. Растениям постоянно «казалось», что они получают свет высокой интенсивности, и таким образом не могли адаптироваться к суточным колебаниям температуры и страдали от ночных заморозков.
При исследовании суточных ритмов у арабидопсис была также показана фотопериодичность работы трёх генов для белков CO, FKF1 и G1. Ген constans участвует в определении времени цветения. Синтез продукта гена — белка CO запускается комплексом из белков FKF1 и G1. В этом комплексе продукт гена FKF1 играет роль фоторецептора. Синтез белка CO запускается через 4 часа после начала освещения и останавливается в темноте. Синтезированный белок за ночь разрушается, и таким образом необходимая для цветения растения концентрация белка достигается только в условиях долгого летнего дня.
Практически все животные приспосабливают свои физиологические и поведенческие процессы к суточным колебаниям абиотических параметров. Примером циркадного ритма у животных является цикл сон-бодрствование. У человека и других животных существуют внутренние часы (часто применяется термин « биологические часы »), которые идут даже в отсутствие внешних стимулов и дают информацию о времени суток. Исследование молекулярно-биологической природы этих часов началось в 1960-х — 1970-х годах . Сеймур Бензер и Рональд Конопка, работавшие в Калифорнийском технологическом институте, обнаружили три мутантные линии дрозофил , циркадные ритмы которых отличались от циркадных ритмов мушек дикого типа . Дальнейший анализ показал, что у мутантов изменения затрагивали аллели одного локуса , который был назван исследователями per (от period).
В отсутствие нормальных сигналов окружающей среды период околосуточной активности у мушек дикого типа составлял 24 часа, у мутантов per-s — 19 часов (short period ), у мутантов per-l — 29 часов (long period ), у мутантов per-0 вообще не наблюдалось никакого ритма. Впоследствии было обнаружено, что продукты генов per есть во многих клетках дрозофил, участвующих в продукции циркадного ритма насекомого. Более того, у мушек дикого типа наблюдаются циркадные колебания в концентрации матричной РНК ( мРНК ) гена per и белка PER (по существующей в молекулярной биологии номенклатуре гены обозначаются строчными, а их белковые продукты — прописными буквами ), в то время как у мушек per-0 , у которых нет циркадного ритма, экспрессия генов не наблюдается.
У млекопитающих главными генами, лежащими в основе циркадианного молекулярного осциллятора супрахиазматического ядра (СХЯ) гипоталамуса , являются гены mPer1 и mPer2 («m» означает «mammalian», то есть period-ген млекопитающих). Экспрессия mPer1 и mPer2 регулируется транскрипционными факторами CLOCK и BMAL1. Гетеромеры CLOCK/BMAL1 связываются с промоторами генов mPer1 и mPer2 , что инициирует их транскрипцию. Образующиеся в результате этого мРНК транслируются в цитоплазме клеток СХЯ в белки mPER1 и mPER2. Эти белки проникают в ядра клеток и, будучи теперь уже связанными с белками mCRY1 и mCRY2, подавляют транскрипцию генов mPer1 и mPer2 , связываясь с CLOCK/BMAL1-белками. Таким образом, по механизму отрицательной обратной связи формируется чередование подъёмов и спадов продукции мРНК, а затем и самих белков mPER1 и mPER2 с фазой, равной приблизительно 24 ч. Этот цикл подстраивается под ритм освещенности .
Существует несколько дополнительных молекулярных циклов, регулирующих циклическую экспрессию генов mPer1 и mPer2 . Белок BMAL1 тоже синтезируется циклически, и его продукция находится в противофазе с ритмом экспрессии генов mPer1 и mPer2 . Транскрипция гена Bmal1 индуцируется белком mPER2 и тормозится белком REV-ERBα. В промоторах генов Cry1 и Cry2 содержится та же нуклеотидная последовательность (Е-box), что и в промоторах генов mPer1 и mPer2 , поэтому транскрипция генов Cry1 и Cry2 позитивно регулируется комплексом CLOCK/BMAL1. То же самое справедливо и для транскрипции гена Rev-Erbα .
Осцилляции, генерируемые на уровне этих генов и белковых продуктов их экспрессии, амплифицируются и распространяются за пределы СХЯ по всему организму. Например, ген вазопрессина , одного из нейромедиаторов СХЯ, также имеет промотор, содержащий E-box, в результате чего за счёт вазопрессина циркадианный сигнал передаётся в другие отделы нервной системы. Другие нейромедиаторные системы, находящиеся под контролем СХЯ — глутамат- и ГАМК-эргические, пептидергические и моноаминергические системы. Также имеется нейрогуморальный путь распространения циркадианного сигнала по всему организму с вовлечением эпифизарного гормона мелатонина .
В зависимости от предмета рассмотрения, биологические часы как понятие, относящееся к чувству времени и ведению суточных ритмов, располагают или в СХЯ, или в эпифизе :261 , или понятие экстраполируется на всю систему :11 .
Первые эксперименты по изоляции людей от таких датчиков времени, как часы и солнечный свет, привели к тому, что у подопытных вырабатывался примерно 25-часовой циркадный ритм. Ошибка эксперимента состояла в том, что участникам разрешалось включать и выключать свет ( торшер с тусклым освещением :241 ) по своему усмотрению. Чрезмерное использование искусственного освещения перед сном приводило к увеличению продолжительности ритма. Последующее более корректно проведённое исследование показало, что период эндогенного циркадного ритма составляет в среднем 24 часа 11 минут . Ещё одно исследование в группе из 157 человек показало следующие результаты:
Циркадный период у женщин оказался несколько короче, чем у мужчин, период менее 24 часов наблюдался у 35 % женщин и у 14 % мужчин .
А. А. Путилов, ссылающийся на данные большинства проведённых экспериментов, указывает среднее значение периода свободно текущего ритма у человека, находящегося в условиях постоянного тусклого освещения, в интервале 23,47—24,64 часа :247 . К. В. Даниленко указывает верхнюю границу интервала 24,78 часа (середина интервала 24,12 часа) .
Эндогенный ход биологических (или внутренних) часов в организме пропорционален периоду суточных ритмов в культуре фибробластов , оценённому на основании экспрессии гена Bmal1 , — это подтверждает, что суточная ритмичность генетически детерминирована .

Одним из наиболее эффективных внешних сигналов («синхронизаторов», или «датчиков времени» — нем. Zeitgeber , англ. time giver ), поддерживающих 24-часовый цикл, является свет. Воздействие света в ранние утренние часы способствует опережению ритма внутренних часов, то есть более раннему пробуждению и последующему за периодом бодрствования засыпанию. Воздействие света в поздние вечерние часы приводит к задержке ритма — более позднему засыпанию и пробуждению. Таким образом световое воздействие ежедневно подстраивает (увлекает, англ. entrainment — увлечение, унос) свободно текущий ритм утром и вечером :247 . В этом процессе участвуют как палочки и колбочки, взаимодействующие с ганглионарными клетками сетчатки глаза, так и особые сетчатки (ipRGC), содержащие пигмент , которые воспринимают синий цвет спектра и напрямую посылают сигнал в СХЯ. Благодаря второму механизму некоторые из слепых людей с полной потерей цветового и светового зрения не испытывают проблем с подстройкой ритма под 24-часовой цикл освещённости :240 .
Среди других возможных синхронизирующих факторов в ряде работ отмечались солнечно-суточные вариации геомагнитного поля :85—87 , достигающие сравнительно больших значений в средних широтах , а также суточные вариации электрического поля атмосферы Земли . Однако пока неизвестно, как эти изменения влияют на происходящие в организме биохимические и биофизические процессы — как действуют приёмники ( рецепторы ) геомагнитных и электрических сигналов (см. Магниторецепция , Электрорецепция ), реагирует ли человек на воздействие всем организмом, отдельными органами или на клеточном уровне. Исследования показывают, что, например, геомагнитные бури вызывают адаптационный стресс, сбивающий циркадный ритм так же, как и резкая смена часовых поясов :85—87 .
У людей в изолированных условиях, например у космонавтов, 24-часовой циркадный ритм поддерживается с помощью освещения . Для возможного полёта людей на Марс проводились исследования по увлечению циркадного ритма человека воздействием освещения с периодом 23,5 часа и 24,65 часа (последнее соответствует периоду марсианских солнечных суток ). Показана возможность такого увлечения воздействием умеренно яркого света в первой или во второй половине запланированного эпизода бодрствования .
В условиях низкой освещённости (до 30 лк ) такие факторы, как стабильный режим сна-бодрствования, приём пищи, положение тела, знание времени суток, неэффективны (по сравнению с циклом свет — темнота) для синхронизации 24-часового циркадного ритма, поэтому фазы суточных ритмов секреции мелатонина и температуры тела сдвигаются в равной степени на более раннее или позднее время, что отражает эндогенный ход центральных биологических часов . Если человек не спит ночью и воспринимает свет или, наоборот, спит днём и не воспринимает свет, то циклические процессы в его организме не получают правильного внешнего сигнала и между ними возникает рассогласование, десинхроноз .
Способность светового воздействия сдвигать фазу внутренних часов как на запаздывание (свет в начале ночи), так и на опережение (свет перед пробуждением) используется в фототерапии , причём воздействие в самом начале или в самом конце ночи приводит к небольшим сдвигам. Чем ближе световое воздействие к середине ночи, тем больше фазовый сдвиг — он может достигать нескольких часов. Световое воздействие в дневном интервале суточного ритма практически не сдвигает его фазу. Для исследования реакции циркадного ритма на внешнее воздействие в практику хронобиологии был введён инструмент под названием « » :244—245 .
Два циркадных ритма с одним и тем же свободно текущим периодом могут по-разному вести себя в зависимости от силы лежащего в их основе колебательного процесса. Генерируемый организмом ритм называют «сильным», если область возможной подстройки его периода достаточно узкая, например, в пределах 23,5—24,5 часа. К группе «сильных» ритмов относятся в первую очередь ритмы температуры тела и секреции мелатонина (самые изученные процессы), а также, например, ритм «бодрость-сонливость». Последний не является простым отражением цикла «сон-бодрствование», относящегося к группе «слабых» ритмов. Для «слабых» ритмов характерна более широкая область подстройки периода. Так, человек, изолированный от внешних сигналов времени, в условиях тусклого освещения может довольно долго жить по навязанному ему режиму сна-бодрствования с периодом 21 или 27 часов (а также 20 или 28 часов ). Под такой режим неспособно подстроиться большинство ритмических процессов, наиболее стабильных в условиях постоянного освещения и наиболее строго контролируемых биологическими часами :242, 248 .
Циркадная система человека имеет индивидуальные отличия. Наиболее ярким их проявлением служит хронотип. Он бывает ранним («жаворонки»), промежуточным («голуби») и поздним («совы»). Люди, относящиеся к раннему хронотипу, ложатся спать и просыпаются в среднем на два часа раньше «сов» и достигают пика интеллектуальной и физической активности в первой половине дня. У людей, относящихся к позднему хронотипу, максимум умственной и физической работоспособности приходится на вторую половину дня. Среди мужчин и двадцатилетней молодежи преобладают «совы», а дети и пожилые люди чаще «жаворонки» .
Как правило (не строго), эндогенная продолжительность циркадного ритма у «жаворонков» меньше 24 часов, а у «сов» больше — у них сон, особенно зимой, уходит на более позднее время :261 .
Некоторые авторы описывают суточные ритмы работы внутренних органов человека . Поскольку статьи с такими сведениями практически или вовсе не цитируются в научной литературе, их ценность сомнительна.
С нарушениями циркадного ритма тесно связаны нарушения сна — джетлаг , расстройство, связанное со сменным графиком работы, и т. п. Также с нарушением циркадного ритма связывают такие нарушения сна, как :