Interested Article - Пластинчатые

- 2021-02-28
- 1
Пласти́нчатые ( лат. Placozoa ) — тип многоклеточных животных , в который включают как минимум 3 вида: Trichoplax adhaerens , Hoilungia hongkongensis и (впрочем, видовое разнообразие Placozoa, по-видимому, гораздо больше). Это маленькие бесцветные организмы , распространённые в спокойных морских водах на небольших глубинах в тропиках и субтропиках ; кроме того, их выращивают в лабораториях по всему миру.
Питаются пластинчатые водорослями и другими микроскопическими пищевыми частицами. Тело Trichoplax представляет собой пластинку диаметром 2—3 мм, состоящую из трёх клеточных слоёв и четырёх (по последним данным, шести) типов клеток. Несмотря на крайнюю простоту организации трихоплакса, в его ядерном геноме обнаружено множество генов , кодирующих белки , необходимые для развития и функционирования высших животных (например, белки, необходимые для развития и работы нервной системы ). Эволюционные связи Trichoplax и природа его простоты (первичность или вторичность) в настоящее время не ясны. Hoilungia hongkongensis по строению и физиологии не отличается от трихоплакса , однако генетический анализ выявил существенные различия между двумя организмами, а также показал наличие между ними репродуктивной изоляции .
История изучения

Впервые трихоплакс был обнаружен австрийским зоологом Францем Шульце в 1883 году на стенках морского аквариума при Зоологическом институте университета Граца , содержавшего образцы растений и животных, найденные в Триестском заливе Адриатического моря . Шульце назвал этот странный организм в виде беловатых пластинок непостоянной формы Trichoplax adhaerens . Название рода образовано от др.-греч. θρίξ (род. п. τριχός ) «волос» и πλάξ «пластинка», а в качестве видового названия взято лат. adhaerens «прилипающий» . При этом Шульце обнаружил, что сторона тела животного, обращённая к субстрату ( вентральная ), покрыта высоким ресничным эпителием, а противоположная ( дорсальная ) сторона покрыта плоским ресничным эпителием (применение термина « эпителий » в случае трихоплакса не совсем корректно; ) .
Вскоре после опубликования работ Шульце итальянский учёный Ф. Монтичелли обнаружил близкую к трихоплаксу форму в морском аквариуме Неаполитанской биологической станции и назвал её Treptoplax reptans . Основным отличием Treptoplax reptans от Trichoplax adhaerens было отсутствие ресничек на дорсальной стороне. Впрочем, после Монтичелли Treptoplax никто не находил, и само существование этого организма вызывает сомнение: Монтичелли мог просто не заметить редко расположенные реснички на дорсальной стороне трихоплакса на гистологических срезах .
Открытие Trichoplax adhaerens вызвало бурный интерес, и вскоре было опубликовано несколько новых работ по его морфологии и биологии. Многие биологи рассматривали трихоплакса как промежуточную форму между одноклеточными и многоклеточными организмами. Некоторые считали его близким к предкам бескишечных турбеллярий ( Acoela ) .
В 1907 году авторитетный немецкий исследователь обнаружил в своём аквариуме трихоплакса именно тогда, когда содержащиеся в нём гидроидные медузы Eleutheria krohni достигли половой зрелости . Это совпадение позволило Крумбаху сделать вывод о том, что Trichoplax adhaerens — это не самостоятельный биологический вид, а аберрантная планула E. krohni . К тому моменту половое размножение у трихоплакса описано не было, и, ввиду авторитетности Крумбаха, его выводы были восприняты без должной критики и начали публиковаться как факт во многих зоологических изданиях (хотя уже в 1912 году Г. Шуботц провёл гистологический анализ строения и клеточного состава T. adhaerens и планул E. krohni и показал, что между данными организмами никакого сходства нет ). Из-за этого в течение последующих 60 лет трихоплакс практически не исследовался; после статьи Ф. Шульце 1914 года в зоологических журналах почти полвека не появлялось ни одной статьи, посвящённой изучению трихоплакса .
Возобновились исследования трихоплакса в 1960-е годы. В 1963 и 1966 годах были опубликованы статьи супругов В. и Г. Куль из Франкфуртского университета , посвящённые передвижению и регенерации трихоплакса. Несколько позже культивированием Trichoplax adhaerens занялись А. В. Иванов и другие советские биологи, изучавшие его строение и биологию .
В 1971 году К. Грелль описал яйцеклетки у Trichoplax adhaerens и изучил его строение на электронно-микроскопическом уровне. Он также разработал методы культивирования трихоплакса в чашках Петри . Существование яйцеклеток у Trichoplax adhaerens послужило доказательством самостоятельности его как биологического вида. Грелль предложил выделить Trichoplax adhaerens в отдельный тип Placozoa. После этого начали активно исследоваться цитологические , молекулярно-биологические , поведенческие и другие особенности трихоплакса . В 2006 и 2008 годах соответственно его митохондриальный и ядерный геномы были секвенированы .
Строение

Уплощённое, лишённое симметрии тело Trichoplax достигает 2—3 мм в диаметре и всего лишь 25 мкм в толщину. В теле Trichoplax выделяют всего 4 типа клеток, организованных в три слоя: верхний и нижний слои клеток, а также разделяющая их соединительная ткань . Наружный слой клеток очень похож на эпителий из-за наличия типичных клеточных контактов между соседними клетками (ленточных десмосом в апикальной области клеток, связанных с пучками актиновых филаментов, а также контактов, похожих на септированные десмосомы), поэтому он больше напоминает эпителий Metazoa , чем губок . Однако его не подстилает , как все настоящие эпителии. Клетки верхней (дорсальной) поверхности плоские, несут по одной ресничке, их ядросодержащие части вдаются вглубь тела. В верхнем слое клеток располагаются так называемые блестящие шары ( нем. Glanzkugeln — термин, введённый Ф. Шульце в 1891 году ), которые представляют собой сильно преломляющие свет липидные тельца и являются остатками дегенерировавших клеток (вероятно, они происходят из клеток срединного волокнистого синцития , мигрировавших в верхний слой). Реснички располагаются на дне структурно оформленного углубления. От базального тельца каждой реснички отходят микротрубочки и пучки поперечно-исчерченных фибрилл, обеспечивающих опору реснички .
Нижняя (вентральная) поверхность обращена к субстрату, покрывающие её клетки представлены безжгутиковыми железистыми клетками и клетками, несущими по одной ресничке. Прочное прикрепление к субстрату (отсюда видовой эпитет adhaerens ) обеспечивается выростами ресничных клеток, напоминающими микроворсинки . Эти клетки высокие и узкие, поэтому реснички располагаются близко друг к другу, благодаря чему на нижней поверхности тела животного образуется плотный ресничный покров, обеспечивающий локомоцию .
Пространство между верхним и нижнем слоями клеток заполнено соединительной тканью. Она представлена волокнистым синцитием, образующим сложную трёхмерную сеть; он залегает в жидкой составляющей, близкой по составу к морской воде из-за более слабых межклеточных контактов, чем у других низших многоклеточных. Её нельзя считать настоящим внеклеточным матриксом , характерным для всех остальных многоклеточных животных, потому что в ней отсутствует коллаген - протеогликан - гликопротеиновый комплекс. Многочисленные ядра мезенхимного волокнистого синцития отделяются друг от друга не мембранами , а внутриклеточными перегородками (септами). Аналогичные септы имеются в синцитиях стеклянных губок и грибов. Впрочем, по последним данным синцитиальное строение сети волокнистых клеток не подтверждается чётко . Ядра этих клеток тетраплоидны , в отличие от диплоидных ядер клеток верхнего и нижнего эпителиев. Их митохондрии связаны в единый комплекс, расположенный вблизи ядра; этот комплекс включает также пузырьки неизвестной природы. Кроме того, волокнистые клетки содержат эндосимбиотические бактерии и, в отличие от клеток верхнего и нижнего слоёв, способны к фагоцитозу . Считается, что волокнистый синцитий сократим, он содержит актин, микротрубочки и, вероятно, миозин . Вероятно, волокнистые клетки функционально соответствуют как мышечным , так и нервным клеткам. Кроме волокнистого синцития, в промежуточном слое залегают половые клетки .
При росте тела трихоплакса митозы происходят во всех трёх клеточных слоях, причём клетки каждого типа, кроме железистых, образуются из себе подобных (железистые клетки образуются от ресничных). у трихоплакса отсутствуют. Эксперименты по регенерации у трихоплакса показали наличие некоторой степени дифференцировки клеток . Так, из изолированной центральной области пластинки и краевого валика шириной около 20 мкм, образованного несколько более мелкими клетками, восстановление цельной пластинки невозможно. При искусственном соединении краевого и центрального фрагментов происходит отторжение лишнего материала, что свидетельствует о существовании определённого баланса между клетками краевой и центральной частей пластинок. Возможно, процесс деления трихоплакса запускается нарушением этого баланса. При несбалансированном росте обоих слоёв клеток проявляются определённые аномалии строения. Так, при недостатке или полном отсутствии дорсального слоя клеток формируются крупные полые шары из вентрального слоя клеток, который проявляет способность к пиноцитозу и изнутри выстилается волокнистыми клетками. Если же отсутствует вентральный эпителий, то формируются сплошные шары из дорсального эпителия, заполненные волокнистым синцитием .
В 2014 году был проведён пересмотр клеточного состава Trichoplax . Согласно новейшим представлениям, в теле трихоплакса выделяют 6 типов соматических клеток : в вентральном слое клеток — ресничные эпителиальные клетки, новоидентифицированные липофильные клетки, заполненные липидными гранулами, и железистые клетки; в промежуточном слое — волокнистые клетки; в дорсальном слое клеток — дорсальные эпителиальные клетки, новоидентифицированные кристаллические клетки. Функциональные и другие особенности типов клеток трихоплакса перечислены в таблице ниже .
| Тип | Описание |
|---|---|
| Вентральные эпителиальные клетки | Мелкие клетки, вытянутые в дорсовентральном направлении, имеют единственную ресничку, поднимающуюся из особого углубления, и множество микроворсинок. Содержат одно крупное (~ 1 мкм) хлопьевидное включение рядом с ядром, плотные гранулы диаметром 200—400 нм вблизи основания, и мелкие пустые везикулы по всей клетке. Вентральные эпителиальные клетки соединены десмосомами в апикальной области, однако характерные для Metazoa плотные контакты и базальная пластинка отсутствуют. Эти клетки можно визуализировать при помощи флуоресцентных антител к мембранно-связанными гуанилаткиназам (MAGUKs), которые часто ассоциированы с клеточными контактами. На долю вентральных эпителиальных клеток приходится 72 % всех клеток в теле трихоплакса. |
| Липофильные клетки | Клетки, лишённые ресничек, разбросаны между вентральными эпителиальными клетками. Содержат крупное (~3 мкм) окружённое мембраной сферическое включение с липидным содержимым, расположенное вблизи вентральной поверхности. Под сканирующим электронными микроскопом липофильные клетки легко отличимы от вентральных эпителиальных клеток благодаря крупным размерам, более глубокому расположению и бугристым очертаниям. Липофильные клетки распределены примерно равномерно в вентральном слое клеток, однако полностью отсутствуют в дорсальном слое. На их долю приходится 11 % клеток в теле трихоплакса, что делает их вторым по многочисленности типом клеток этого организма. |
| Железистые клетки | Клетки вентрального эпителия, как предполагается, выполняют секреторную функцию. Содержат многочисленные гранулы размером 200—500 нм , которые в некоторых клетках тёмные, а в некоторых — бледные; гранулы разбросаны по всему телу клетки. Железистые клетки наиболее многочисленны в краевой области пластинки (в районе 20 мкм от края), по направлению к центру их численность уменьшается. |
| Волокнистые клетки | Клетки соединительнотканного слоя, имеют удивительно ровные края. Отростки волокнистых клеток контактируют с дорсальными поверхностями клеток вентрального слоя и проникают между телами этих клеток, кроме того, они контактируют с дорсальным слоем, другими волокнистыми клетками и липофильными клетками. Впрочем, настоящих клеточных контактов они не образуют. Синцитиальная структура волокнистых клеток не находит чёткого подтверждения. Для волокнистых клеток характерно наличие кластера митохондрий, в который проникают более бледные тельца, палочковидные включения (возможно, эндосимбиотические бактерии), окружённые шероховатым эндоплазматическим ретикулумом , а также крупной конкрементной вакуоли, содержащие остатки водорослей, съеденных организмом. Волокнистые клетки можно визуализировать методом иммунофлуоресценции при помощи антител к предполагаемому кадгерину трихоплакса, а также рецептору . На долю волокнистых клеток приходится 4,4 % всех клеток животного. |
| Дорсальные эпителиальные клетки | Подобно клеткам вентрального эпителия, клетки дорсального эпителия соединены межклеточными контактами и имеют реснички и микроворсинки, хотя микроворсинок обычно меньше, чем у клеток вентрального эпителия. Они содержат многочисленные плотные эллиптические гранулы ~0,5 мкм в диаметре. Эти клетки можно визуализировать при помощи антител к MAGUKs и фибриллярному актину. Форма клеток может изменяться от сферической до эллиптической, что соответствует изменениям формы тела животного. Дорсальные эпителиальные клетки составляют 9 % клеток трихоплакса. |
| Кристаллические клетки | Кристаллические клетки содержат ромбоидные кристаллы ~ 2 мкм в диаметре, чашевидное ядро, приуроченное к одной стороне клетки, и две центрально расположенные митохондрии, фланкирующие кристалл. Цитоплазма кристаллических клеток лишена включений. Кристаллические клетки лежат под дорсальным эпителием и соседствуют с волокнистыми клетками и липофильными клетками. Они лежат дорсальнее железистых клеток и не имеют входа на поверхность. Точные функции кристаллических клеток неизвестны, но они могут выполнять роль статоцистов или фоторецепторов . Они составляют менее 0,2 % всех клеток в теле трихоплакса. |
Иногда в старых культурах появляются нежизнеспособные сферические формы трихоплакса. При образовании этих сферических форм дорсальный слой клеток утрачивает контакт с волокнистым синцитием, и слой соединительной ткани наполняется жидкостью. Если при этом вентральный слой клеток отделяется от субстрата, то сферическая форма трихоплакса может некоторое время плавать в культуре как маленький шарик. Выделяют два типа сферических форм: полые , наружная стенка которых представлена ресничным вентральным эпителием, а внутрь полости вдаются волокнистые клетки, причём в полости могут также содержаться ресничные дорсальные клетки; плотные , наружная стенка которых образована дорсальным эпителием, а внутренняя часть плотно набита волокнистыми клетками (кроме того, в ней может находиться компартмент вентральных клеток) .
Биология
По способу перемещения трихоплакс напоминает амёбу : он медленно скользит по субстрату, постоянно меняя очертания за счёт наличия связей между клетками верхних и нижних слоёв. Постоянные передний и задний концы тела не выражены, поэтому трихоплакс может изменять направление движения, не поворачиваясь. При попытке начать ползти в двух противоположных направлениях одновременно тело трихоплакса может разорваться на две части. Сужение и разрыв перетяжки при делении также происходит за счёт контактов между клетками в верхнем и нижнем слоях .

Trichoplax adhaerens питается водорослями и другими пищевыми частицами, находящимися на субстрате. Пищеварение внеклеточное, происходит вне тела трихоплакса, между его вентральной поверхностью и субстратом. Во время питания трихоплакс изгибается, приподнимая над субстратом центральную часть, и в образовавшемся замкнутом кармане происходит переваривание пищи; этот процесс иногда называют «временной гаструляцией », причём вентральный эпителий трихоплакса функционально соответствует стрекающих . Пищеварительные ферменты , вероятно, выделяются железистыми клетками. Продукты пищеварения поглощаются ресничными клетками, которые могут образовывать эндоцитозные окаймлённые везикулы. По-видимому, маленькое плоское тело делает возможным транспорт путём обычной диффузии , поэтому необходимости в циркуляторной системе у трихоплакса нет .
Волокнистые клетки содержат конкрементные (пищеварительные) вакуоли , в которых можно обнаружить пищевые частицы на разных стадиях переваривания. Вероятно, отростки волокнистых клеток проникают сквозь верхний слой клеток и там фагоцитируют пищевые частицы .
В 2015 году была опубликована статья, в которой сообщалось об интересной детали пищевого поведения трихоплакса. Оказалось, что чем больше водорослей оказалось в пищевом кармане трихоплакса, тем больше липофильных клеток выделяют свой секрет наружу, заставляя клетки водорослей лизироваться . Кроме того, разряжаются только те липофильные клетки, которые находятся на расстоянии 10−20 мкм от пищевой частицы. Механизмы обеспечения такой точной регуляции неизвестны .
Имеются сведения о существовании группового поведения у трихоплакса. По некоторым данным, при обитании в больших аквариумах, моделирующих естественные условия, трихоплакс существует в виде скоплений, развитие которых проходит через определённые стадии .
Размножение и развитие
Trichoplax adhaerens размножается преимущественно бесполым путём ( фрагментацией или почкованием ). При фрагментации тело трихоплакса с помощью образующейся перетяжки разделяется на две примерно равные половинки. Процесс фрагментации может занимать несколько часов, причём между расходящимися особями длительное время сохраняется тонкий многоклеточный мостик .
При почковании округлые почки (бродяжки) диаметром 40—60 мкм образуются на дорсальной поверхности, они содержат все типы клеток, в том числе вентральные ресничные клетки и соединительную ткань. Формирование почек происходит следующим образом. Зачаток почки формируется во внутреннем слое тела из клеток, которые выселяются внутрь из верхнего и нижнего слоёв, теряя при этом реснички. В дальнейшем эти клетки образуют вентральный эпителий дочерней особи. До отделения почки эти клетки формируют реснички, направленные внутрь полости и, таким образом, меняют полярность на противоположную. По мере роста почка образует выпячивание на дорсальной стороне тела и в конце концов отшнуровывается от неё вместе с частью дорсального слоя клеток. Процесс почкования длится около 24 часов. Отделившись, покрытые ресничками почки уплывают, плавают около недели и оседают на дно. На обращённой к субстрату стороне образуется отверстие, которое постепенно расширяется, так что бродяжка становится чашеобразной, а после распластывается по субстрату и прикрепляется к нему вентральным эпителием. От краевого валика могут отделяться более мелкие почки, образованные клетками дорсального и вентрального слоёв и заполненные волокнистыми клетками. Такие более мелкие почки не способны к плаванию в толще воды и выполнению расселительной функции, они постепенно уплощаются и превращаются в молодых особей .
Достоверного описания полового процесса у трихоплакса пока нет, однако в лабораторных условиях у него были описаны яйца, а на ультраструктурном уровне — и спермии . Существование полового процесса также подтверждается распределением однонуклеотидных полиморфизмов . Яйцеклетки образуются из клеток нижнего слоя, которые дедифференцируются и погружаются в слой соединительной ткани. Клетки волокнистого синцития играют роль кормящих, часть их отростков вместе с эндосимбиотическими бактериями фагоцитируют яйцеклетки. Половые клетки у трихоплакса образуются нерегулярно; к факторам, запускающим половое размножение, можно отнести увеличение плотности популяции , нехватку пищи и повышение температуры воды до 23 °C и более. И яйцеклетки, и спермии могут формироваться на одной особи. Характер мейоза неизвестен. Яйцеклетка оплодотворяется по достижении размера в 70—120 мкм. После этого яйцо покрывается так называемой защитной «оболочкой оплодотворения» и претерпевает полное равномерное дробление . Зародыш развивается внутри материнского организма, пока последний полностью не разрушится и новый организм не выйдет наружу. В лабораторных условиях зародыши разрушаются на стадии 64 бластомеров по неизвестным причинам (материнская особь тоже гибнет), так что эмбриональное развитие трихоплакса не изучено. Возможной причиной гибели зародыша может быть неконтролируемая репликация ДНК , при которой не происходит перехода клеток из S-фазы в G 2 -фазу .
Молекулярная биология
Хромосомный набор Trichoplax adhaerens 2n = 12 . Три пары представлены двуплечными мета- или субметацентрическими хромосомами , в то время как три другие пары хромосом имеют меньший размер и, возможно, являются акроцентрическими. Абсолютный размер хромосом не превышает 2—3 мкм . Диплоидные эпителиальные клетки содержат всего 0,08 пг ДНК , что меньше аналогичного значения у любого представителя Metazoa .
Секвенированию ядерного генома Trichoplax adhaerens предшествовало секвенирование его митохондриального генома , осуществлённое в 2006 году. Результаты были неожиданными: оказалось, что митохондриальный геном T. adhaerens имеет наибольший размер среди всех Metazoa и содержит более 43 тыс. пар нуклеотидов (в то время как размеры митогеномов других Metazoa обычно лежат в пределах от 15 до 24 тыс. пар нуклеотидов). К отличительным его особенностям следует отнести наличие многочисленных спейсеров между участками транскрибируемой ДНК, нескольких интронов , а также ORF с пока невыясненными функциями (всё это нетипично для других Metazoa), а участки ДНК, кодирующие белки, в среднем на 10 % больше, чем у других животных. Перечисленные особенности митохондриального генома трихоплакса следует считать признаками, унаследованными от общего предка Metazoa, поскольку среди опистоконтов , не относящихся к Metazoa, данные особенности — не редкость. Так, у хоанофлагеллаты Monosiga brevicollis митохондриальный геном содержит примерно 76,5 тыс. пар нуклеотидов, имея большие области, занятые спейсерами, и несколько интронов (а митохондриальный геном аскомицета состоит из 94—100 тыс. пар нуклеотидов ) .
С учётом того, что среди пластинчатых ранее было выявлено наличие значительного генетического разнообразия, нашедшего отражение в выделении нескольких достаточно обособленных клад , исследователи в 2007 году секвенировали митохондриальные геномы ещё у трёх лабораторных образцов, полученных на территории Белиза . Среди них образец под условным обозначением BZ2423 представлял кладу II, BZ10101 — кладу III, а BZ49 — кладу V (сам T. adhaerens принадлежит к кладе I). Анализ этих геномов показал, что они разделяют ранее выявленные у митохондриального генома T. adhaerens особенности: во всех четырёх геномах имеются многочисленные спейсеры, присутствуют несколько ORF с неизвестными функциями, интроны, причём выделен общий набор генов: 12 генов дыхательной цепи , 24 гена тРНК , 2 гена рРНК ( rnS и rnL ), а гены at8 и atp9 не обнаружены. В то же время наблюдалось значительное различие в размерах генома: у образца BZ49 он содержал 37,2 тыс. пар нуклеотидов, у BZ2423 — 36,7 тыс. пар, а у BZ10101 — 32,7 тыс. пар. Доля кодирующих и структурных последовательностей нуклеотидов варьировала от 55 % у T. adhaerens до 67 % у BZ49. Средняя длина спейсера менялась от 101 пары у BZ10101 и 105 пар у BZ49 до 154 пар у BZ2423 и 209 пар у T. adhaerens .
Ядерный геном трихоплакса был секвенирован в 2008 году. Он содержит около 98 млн пар нуклеотидов и 11514 белоккодирующих генов, из которых около 87 % обнаруживают значительное сходство с соответствующими генами других животных. Большая часть (около 83 %) из около 7800 , консервативных у актиний и двусторонне-симметричных животных, имеют гомологи у трихоплакса. Плотность интронов у трихоплакса сопоставима с таковой у позвоночных и актиний. В ядерном геноме Trichoplax сохранились многие древние интроны, утраченные в других линиях животных, и для 82 % интронов человека находятся соответствующие интроны с тем же расположением в генах трихоплакса . Интересно, что некоторые гены трихоплакс приобрёл от своего бактериального грамотрицательного эндосимбионта ( риккетсии ) . Ядерный геном трихоплакса довольно мал по сравнению с геномами Metazoa, однако это вряд ли можно объяснить вторичным упрощением: об отсутствии вторичного упрощения свидетельствует сохранение древних интронов и порядка генов ( синтении ) .
Несмотря на относительную простоту строения, генетический материал Trichoplax adhaerens содержит множество генов, присущих животным с более высоким уровнем организации . Например, вторичная структура молекулы 16S рРНК у трихоплакса существенно сложнее, чем у любых представителей типа стрекающих ( Cnidaria ) . Ядерная ДНК трихоплакса содержит гены белков (в особенности, транскрипционных факторов ), необходимых для дифференцировки различных типов клеток, развития и функционирования нервной системы, обособления клеток зародышевой линии и других аспектов развития двусторонне-симметричного организма. В ней имеются гены Hox и и другие гомеобоксные гены: , Brachyury и . У трихоплакса также есть гены, кодирующие белки сигнальных путей - и Wnt , которые чрезвычайно важны для определения осей тела в ходе развития двусторонне-симметричного организма .
Кроме того, у трихоплакса обнаружены гены, кодирующие белки базальной пластинки, молекулы, необходимые для межклеточной адгезии и прикрепления клеток к внеклеточному матриксу. Трихоплакс имеет также гены, критичные для работы нервной системы — такие, как гены, кодирующие потенциал-зависимые натриевые , калиевые и кальциевые каналы , почти полный набор синаптических , белки систем синтеза и транспорта моноаминов , множество ионотропных и G-связанных рецепторов ( GPCR ), факторы поддержания аксонов и миграции нейронов. Установлено, что у трихоплакса имеются зрительные пигменты — опсины . Имеются доказательства наличия у трихоплакса генов инсулина и инсулиновых рецепторов , а также генов различных регуляторных пептидов и их рецепторов, что свидетельствует о наличии зачатков гормональной регуляции у трихоплакса . У Trichoplax adhaerens обнаружен ген предшественника главного комплекса гистосовместимости (прото-МНС), который необходим для функционирования иммунной системы позвоночных; это рассматривают как аргумент в пользу предположения о раннем возникновении прото-МНС в эволюции животных. У трихоплакса прото-МНС участвует в защитных процессах (противо вирусный иммунитет, ответ на стресс, убиквитиново - протеасомный путь расщепления белков) .
У трихоплакса обнаружен особый мембранноактивный антимикробный пептид — трихоплаксин, который может быть использован для разработки нового ряда антимикробных препаратов широкого спектра действия .
Разнообразие, распространение и экология

Долгое время вид Trichoplax adhaerens считался единственным представителем типа Placozoa, поскольку в световой микроскоп все пластинчатые из разных частей мира выглядят одинаково. Ряд недавних исследований опроверг это представление. Выяснилось, что среди пластинчатых имеет место значительное генетическое разнообразие, и по состоянию на 2015 год таксон Trichoplax adhaerens в действительности включает 19 видов (пока они именуются гаплотипами ), составляющих по меньшей мере 7 хорошо обособленных клад . Описание отдельных видов в случае трихоплакса представляется чрезвычайно сложным из-за однообразной морфологии этих животных .
В 2018 году был выделен новый вид пластинчатых — Hoilungia hongkongensis . Было проведено секвенирование генома этого организма, и его геном был сравнён с геномом T. adhaerens . Оказалось, что у H. hongkongensis имеется много инверсий и транслокаций , отсутствующих в геноме T. adhaerens , что указывает на глубокое разделение между двумя видами. Анализ дивергенции последовательностей также указал на неожиданно большое эволюционное расстояние между двумя видами пластинчатых. Было также показано, что ключевой механизм видообразования у пластинчатых (то есть разделения вида на два новых) — дупликации генов .
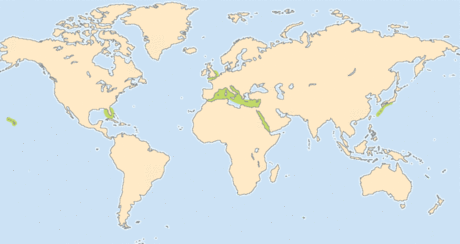
Об экологии и биогеографии трихоплаксов известно очень немного. Пластинчатые обнаруживаются только в спокойных прибрежных водах и избегают глубоководий, а также вод с сильными течениями. Они были найдены и в стоячих водах на глубине до 20 м. Считается, что основными факторами, ограничивающими распространение трихоплаксов, являются температура воды и её солёность. Пластинчатые были обнаружены в тропических и субстропических водах Индийского , Тихого и Атлантического океанов, а именно, вблизи Бермудских островов , в Карибском море , у побережья Восточной Австралии , в Большом Барьерном рифе , около острова Гуам , Гавайев , Японии , в Средиземном море , у островов Палау , Папуа-Новой Гвинеи , в Красном море , вблизи Вьетнама и Восточного Самоа . Члены некоторых клад распространены очень широко, в то время как ареал других клад более ограничен .
Пластинчатые предпочитают тёплую воду с температурой до 32 °С, причём иногда они обнаруживались в воде с температурой чуть выше 10 °С. Чаще всего пластинчатые встречаются в местах со спокойными водами на твёрдых субстратах (камнях, коралловых рифах , корнях мангровых деревьев ). Разнообразие сред обитания пластинчатых может лежать в основе их большого видового разнообразия. Пластинчатые могут приспосабливаться к мутным водам, а также низкой солёности воды, которая, например, наблюдается в мангровых зарослях .
Численность трихоплаксов зависит от времени года. Как правило, пластинчатые наиболее многочисленны в тёплое время года (с июня по октябрь в субтропическом и умеренном поясах Северного полушария ) .
Наблюдения за связями трихоплаксов с другими живыми организмами показывают, что они часто ассоциированы с сидячими фильтраторами, например, сидячими инфузориями и многощетинковыми червями . Из-за малых размеров и бентосного образа жизни пластинчатые должны часто становиться жертвами хищников, однако пока известен лишь один случай поедания трихоплакса. Описаны случаи, когда потенциальные хищники (например, брюхоногие моллюски или стрекающие) быстро отскакивали после контакта с трихоплаксом, что свидетельствует о возможном существовании химических механизмов защиты у этих животных. Может быть, они опосредуются блестящими шарами, расположенными в верхнем слое клеток .
Для сбора пластинчатых из естественных местообитаний существуют два основных метода. Первый из них подразумевает сбор камней, обломков коралловых рифов и других твёрдых объектов с глубины до 5 м. Во втором методе используются обычные предметные стёкла, которые помещаются в пластиковый пакет, открытый с двух сторон, так что через него возможно течение морской воды. Пакеты со стёклами погружаются на глубину от 2 до 20 м. Вскоре стёкла покрываются налётом из водорослей, которые служат кормом для пластинчатых . В лабораторных условиях трихоплаксов кормят одноклеточными водорослями и Chlorella .
Эволюция и классификация

Относительно простоты строения Trichoplax нет единого мнения: неизвестно, является ли она первичной, или же она является результатом вторичного упрощения. Существуют даже предположения, что трихоплакс представляет собой прогенетическую личинку какого-то полностью исчезнувшего таксона . Согласно данным анализа митохондриальной ДНК , Placozoa были первой группой, отделившейся вблизи основания эволюционного древа животных. Однако анализ нескольких ядерных генов дал противоречивые результаты. Согласно одним данным, пластинчатых следует рассматривать как базальную группу Metazoa, согласно другим — как вторую ветвь, отделившуюся от древа животных после отделения губок, согласно третьим — как сестринскую группу Bilateria . Анализ 18S рРНК и генов Hox/ParaHox свидетельствует в пользу монофилии группы, включающей таксоны Placozoa, Cnidaria и Bilateria и названной . Недавние палеонтологические находки, проливающие свет на модель питания крупных мягкотелых эдиакарских животных (таких, как Dickinsonia ), позволяют заключить, что эти организмы близки к современным пластинчатым, хотя достоверных ископаемых остатков собственно пластинчатых не найдено .
Многие учёные рассматривали Trichoplax как промежуточное звено между одноклеточными и многоклеточными животными. Под влиянием первых сведений о трихоплаксе О. Бючли выдвинул гипотезу происхождения многоклеточных от плакулы — гипотетической двуслойной колонии. Её нижний слой был обращён к субстрату и выполнял трофическую функцию (будущая энтодерма ), а верхний выполнял защитную функцию и дал начало эктодерме . С этой гипотезой хорошо согласуется пищеварительная роль вентрального эпителия Trichoplax . Однако способность волокнистых клеток к трансэпителиальному фагоцитозу больше соответствует модели фагоцителлы , предложенной И. И. Мечниковым .
Название типа Placozoa предложил К. Грелль в 1971 году. В противовес этому А. В. Иванов , отдававший предпочтение не модели плакулы, а модели фагоцителлы, предложил выделить тип Phagocytellozoa с единственным родом Trichoplax . Несколько позже Иванов (хотя и с замечаниями по поводу «неудачности» названия) признал тип Placozoa, но выделил в системе Metazoa надраздел Phagocytellozoa, включающий единственный тип Placozoa (русский эквивалент названия — Пластинчатые ) . Впрочем, позднейшие исследования выявили, что трихоплакс и модель фагоцителлы не имеют между собой ничего общего . Иногда род Trichoplax выделяют в отдельное семейство Trichoplacidae в составе типа Placozoa .
В 2019 году был описан новый вид пластинчатых — Polyplacotoma mediterranea , имеющий значительные отличия во внешнем строении и в митохондриальном геноме .
Ниже приводится краткий обзор существующих в данный момент гипотез относительно эволюционного положения и происхождения пластинчатых.
Функционально-морфологическая гипотеза
Из-за своего простого строения Placozoa часто рассматривались как модельный организм, иллюстрирующий переход от одноклеточных организмов к многоклеточным животным (Metazoa), а потому Placozoa можно считать сестринской группой по отношению ко всем другим метазоям :
| Metazoa |
|
||||||||||||
Эта идея, остающаяся в настоящее время чисто умозрительной, получила название функционально-морфологической гипотезы. Она предполагает, что все или большинство животных произошли от галлертоида (от нем. Gallert ‘желе, студень’) — свободноживущего морского пелагического сферического организма. Он состоял из единственного слоя ресничных клеток, поддерживаемого тонким неклеточным разделительным слоем — базальной пластинкой. Внутренняя часть сферы была заполнена способными к сокращениям волокнистыми клетками и желеобразным внеклеточным матриксом .
Современные Placozoa и другие животные якобы произошли от этой предковой формы, пройдя один из двух путей. Первый путь — впячивание эпителия — привёл к развитию внутренней системы каналов, и такой модифицированный галлертоид с внутренней системой каналов дал начало губкам, стрекающим и гребневикам . Второй путь включал переход от пелагического образа жизни к бентосному . В отличие от морской воды, где пища, потенциальные хищники и половые партнёры могут встретиться со всех сторон, при жизни вблизи субстрата возникает различие между стороной, обращённой к субстрату, и стороной, обращённой к открытому водному пространству. В этих условиях уплощение тела имеет селективное преимущество, и, в самом деле, уплощённое тело присуще многим бентосным животным. Согласно функционально-морфологической модели, пластинчатые и, возможно, некоторые другие группы, известные только из ископаемых остатков, произошли от такой бентосной формы жизни, названной плакулоидом . По этой модели, дорсальный и вентральный слои клеток трихоплакса являются аналогами , но не гомологами эктодермы и энтодермы эуметазоев .
В случае истинности вышеописанной модели Trichoplax adhaerens оказывался бы древнейшей ветвью многоклеточных организмов и реликтом эдиакарской (или даже пре-эдиакарской) фауны. Из-за отсутствия внеклеточного матрикса и базальной пластинки развитие этих животных, очень успешных в своей экологической нише, было, разумеется, ограничено, что объясняет низкую скорость эволюции пластинчатых и их почти полную неизменность в ходе эволюции .
|
Простой пелагический
галлертоид |
|||||||||||||||||||||||||||||||||||
|
Уплощённый бентосный
плакулоид |
Галлертоид с системой
внутренних каналов |
||||||||||||||||||||||||||||||||||
|
Пре-эдиакарские следы
ползающих организмов (?) |
Placozoa |
Эдиакарская
фауна (?) |
Porifera | Cnidaria | Ctenophora | ||||||||||||||||||||||||||||||
Функционально-морфологическая гипотеза подтверждается анализом митохондриальной ДНК , однако не находит должного подтверждения в статистическом анализе генома трихоплакса по сравнению с геномами других животных .
Гипотеза Epitheliozoa
Согласно гипотезе Epitheliozoa, основанной на чисто морфологических особенностях, Placozoa являются ближайшими родственниками животных с настоящими тканями (Eumetazoa). Образуемый ими таксон (Epitheliozoa), оказывается сестринской группой по отношению к губкам :
| Metazoa |
|
||||||||||||
Главным подтверждением этой гипотезы является наличие особых клеточных контактов — ленточных десмосом, которые имеются у Placozoa и всех остальных животных, за исключением губок. Кроме того, Trichoplax adhaerens объединяет с эуметазоями наличие железистых клеток на вентральной стороне. В рамках этой гипотезы возможным сценарием происхождения моноцилиарных клеток, похожих на эпителий (эпителиоидных) тканей Trichoplax adhaerens , является редукция воротничка хоаноцитов у группы губок, которая утратила способность к фильтрационному питанию и стала предковой группой пластинчатых. Эпителиоидная ткань трихоплакса впоследствии дала начало эпителию эуметазоев .
Согласно модели Epitheliozoa, дорсальный и вентральный слои клеток трихоплакса являются гомологами эктодермы и энтодермы, а слой волокнистых клеток дал начало соединительным тканям других животных. Статистический анализ генома трихоплакса по сравнению с геномами других животных говорит в пользу этой модели .
Гипотеза Eumetazoa
Гипотеза Eumetazoa, базирующаяся в основном на данных молекулярной генетики, рассматривает пластинчатых как крайне упрощённых эуметазоев . Согласно этой гипотезе, трихоплакс произошёл от гораздо более сложно устроенных животных, которые имели мышечную и нервную ткань . Два этих типа тканей, а также базальная пластинка и эпителий были утрачены в ходе последующего радикального упрощения. Было предпринято много попыток определения сестринской группы Placozoa: согласно одним данным, они являются ближайшими родственниками стрекающих, по другим — сестринской группой гребневиков, а иногда их помещают в группу двусторонне-симметричных ( Bilateria ) :
| Metazoa |
|
||||||||||||||||||||||||
Главным недостатком этой гипотезы является то, что она практически не учитывает морфологические особенности пластинчатых. Крайняя степень упрощения, которая, согласно этой модели, имеет место у Placozoa, известна только у животных, ведущих паразитический образ жизни и, по-видимому, маловероятна у свободноживущих видов, каким является Trichoplax adhaerens . Статистический анализ генома трихоплакса по сравнению с геномами других животных отвергает гипотезу .
Гипотеза Diploblasta
Иную картину филогенетических взаимоотношений между основными подразделениями Metazoa дают результаты комбинированного анализа, основанного на совместном использовании морфологических данных, вторичной структуры митохондриальных генов рРНК и нуклеотидных последовательностей из митохондриального и ядерного геномов. На основании этих результатов пластинчатые группируются вместе с губками, гребневиками и стрекающими в кладу Diploblasta, сестринскую к Bilateria :
| Metazoa |
|
||||||||||||||||||||||||
Объединение упомянутых четырёх типов (Placozoa, Porifera, Ctenophora, Cnidaria) в единый таксон высшего ранга (« подцарство Radiata») предложил Т. Кавалье-Смит в своей шестицарственной системе органического мира 1998 года (впрочем, он не настаивал на монофилии данного таксона) . В системе царства Metazoa, предложенной в 2015 году Майклом Руджеро с соавторами и входящей как составная часть в представленную ими макросистему живущих организмов, это царство включает два подцарства: не получившее названия (соответствует группе Diploblasta на приведённой кладограмме) и Bilateria. Данный вариант классификации пригоден и при принятии любой из трёх предыдущих филогенетических гипотез, но тогда указанное безымянное подцарство будет представлять собой уже не кладу, а парафилетическую группу .
Примечания
- DuBuc Timothy Q , Ryan Joseph , Martindale Mark Q. (англ.) // Molecular Biology and Evolution. — 2019. — 6 February. — ISSN . — doi : .
- ↑ Eitel M. , Francis W. R. , Varoqueaux F. , Daraspe J. , Osigus H. J. , Krebs S. , Vargas S. , Blum H. , Williams G. A. , Schierwater B. , Wörheide G. (англ.) // PLoS Biology. — 2018. — July ( vol. 16 , no. 7 ). — P. e2005359—2005359 . — doi : . — .
- Eitel M. , Francis W. R. , Varoqueaux F. , Daraspe J. , Osigus H. J. , Krebs S. , Vargas S. , Blum H. , Williams G. A. , Schierwater B. , Wörheide G. (англ.) // PLoS Biology. — 2018. — September ( vol. 16 , no. 9 ). — P. e3000032—3000032 . — doi : . — .
- ↑ Eitel M., Osigus H. J., DeSalle R., Schierwater B. // PLoS One . — 2013. — Vol. 8, no. 4. — P. e57131. — doi : . — .
- , с. 9.
- ↑ , с. 5—6.
- ↑ Syed T., Schierwater B. // Vie et Milieu. — 2002. — Vol. 52, no. 4. — P. 177—187. 17 июля 2021 года.
- , с. 6—7.
- , с. 11.
- , с. 8.
- , с. 12, 14.
- , p. 294.
- , с. 8—9.
- , p. 310.
- ↑ Srivastava M., Begovic E., Chapman J., Putnam N. H., Hellsten U., Kawashima T., Kuo A., Mitros T., Salamov A., Carpenter M. L., Signorovitch A. Y., Moreno M. A., Kamm K., Grimwood J., Schmutz J., Shapiro H., Grigoriev I. V., Buss L. W., Schierwater B., Dellaporta S. L., Rokhsar D. S. // Nature . — 2008. — Vol. 454, no. 7207. — P. 955—960. — doi : . — .
- ↑ Nikitin M. // General and Comparative Endocrinology. — 2015. — Vol. 212. — P. 145—155. — doi : . — .
- ↑ , с. 177.
- , с. 131—132.
- ↑ , с. 131.
- ↑ Smith C. L., Varoqueaux F., Kittelmann M., Azzam R. N., Cooper B., Winters C. A., Eitel M., Fasshauer D., Reese T. S. // Current Biology. — 2014. — Vol. 24, no. 14. — P. 1565—1572. — doi : . — .
- ↑ , с. 132.
- , с. 133.
- Thiemann, M. Spherical forms of Trichoplax adhaerens (Placozoa) / M. Thiemann, A. Ruthmann // Zoomorphology. — 1990. — Vol. 110, no. 1. — P. 37—45. — ISSN . — doi : .
- , с. 177, 179.
- ↑ , с. 179.
- , с. 132—133.
- Smith C. L., Pivovarova N., Reese T. S. // PLoS One . — 2015. — Vol. 10, no. 9. — P. e0136098. — doi : . — .
- , с. 33.
- , с. 37.
- ↑ , с. 134.
- Eitel M., Guidi L., Hadrys H., Balsamo M., Schierwater B. // PLoS One . — 2011. — Vol. 6, no. 5. — P. e19639. — doi : . — .
- Birstein, V. J. // Biologisches Zentralblatt. — 1989. — Vol. 108, no. 1. — С. 63—67 . 5 марта 2016 года.
- Cummings D. J. , McNally K. L. , Domenico J. M. , Matsuura E. T. (англ.) // Current genetics. — 1990. — Vol. 17, no. 5 . — P. 375—402. — .
- ↑ , p. 299, 310.
- ↑ Dellaporta S. L., Xu A., Sagasser S., Jakob W., Moreno M. A, Buss L. W., Schierwater B. // Proc. Nat. Acad. Sci. USA . — 2006. — Vol. 103, no. 23. — P. 8751—8756. — doi : . — .
- ↑ Signorovitch A. Y., Buss L. W., Dellaporta S. L. // . — 2007. — Vol. 3, no. 1. — P. e13. — doi : . — .
- Driscoll T., Gillespie J. J., Nordberg E. K., Azad A. F., Sobral B. W. // Genome Biology and Evolution. — 2013. — Vol. 5, no. 4. — P. 621—645. — doi : . — .
- ↑ Schierwater B., de Jong D., DeSalle R. // The International Journal of Biochemistry & Cell Biology. — 2009. — Vol. 41, no. 2. — P. 370—379. — doi : . — .
- , p. 296.
- Глаголев, Сергей. . // Сайт Элементы.ru (4 января 2016). Дата обращения: 11 марта 2016. 12 марта 2016 года.
- Suurväli J., Jouneau L., Thépot D., Grusea S., Pontarotti P., Du Pasquier L., Rüütel Boudinot S., Boudinot P. // Journal of Immunology. — 2014. — Vol. 193, no. 6. — P. 2891—2901. — doi : . — .
- Simunić J., Petrov D., Bouceba T., Kamech N., Benincasa M., Juretić D. // Biochimica et Biophysica Acta — Biomembranes. — 2014. — Vol. 1838, no. 5. — P. 1430—1438. — doi : . — .
- ↑ Paknia O., Schierwater B. // PLoS One . — 2015. — Vol. 10, no. 11. — P. e0140162. — doi : . — .
- ↑ . // University of California Museum of Paleontology. Дата обращения: 11 марта 2016. 25 марта 2018 года.
- Sperling E. A., Vinther J. // Evolution & Development. — 2010. — Vol. 12, no. 2. — P. 201—209. — doi : . — .
- Иванов, А. В. . // Жизнь животных : в 7 т. / под ред. Ю. И. Полянского . — 2-е изд. — М. : Просвещение , 1987. — Т. 1. — С. 124—127.
- , с. 45—46.
- Eitel M. . // World Register of Marine Species. Дата обращения: 11 марта 2016. 12 марта 2016 года.
- Osigus H.-J., Rolfes S. , Herzog R., Kamm K., Schierwater B. Polyplacotoma mediterranea is a new ramified placozoan species (англ.) // Current Biology. — 2019. — Vol. 29, no. 5 . — P. R148—R149. — ISSN . — doi : . — .
- ↑ , с. 47—48.
- Schierwater B., Eitel M., Jakob W., Osigus H.-J., Hadrys H., Dellaporta S. L., Kolokotronis S. O., DeSalle R. // PLoS Biology . — 2009. — Vol. 7, no. 1. — P. e20. — doi : . — .
- , p. 301, 303.
- Cavalier-Smith T. // Biological Reviews of the Cambridge Philosophical Society. — 1998. — Vol. 73, no. 3. — P. 203—266. — doi : . — .
- Ruggiero Michael A. , Gordon Dennis P. , Orrell Thomas M. , Bailly Nicolas , Bourgoin Thierry , Brusca Richard C. , Cavalier-Smith Thomas , Guiry Michael D. , Kirk Paul M. (англ.) // PLOS ONE. — 2015. — 29 April ( vol. 10 , no. 4 ). — P. e0119248 . — ISSN . — doi : .
- Ruggiero M. A. , Gordon D. P. , Orrell T. M. , Bailly N. , Bourgoin T. , Brusca R. C. , Cavalier-Smith T. , Guiry M. D. , Kirk P. M. (англ.) // PLOS ONE. — 2015. — 11 June ( vol. 10 , no. 6 ). — P. e0130114 . — ISSN . — doi : .
Литература
- Зоология беспозвоночных : в 2 т. / под ред. В. Вестхайде и Р. Ригера . — М. : Товарищество научных изданий КМК, 2008. — Т. 1: от простейших до моллюсков и артропод. — 512, [8] с. — ISBN 978-5-87317-491-1 .
- Малахов В. В. . Загадочные группы морских беспозвоночных. — М. : Издательство Московского университета, 1990. — 144 с. — ISBN 5-211-00921-5 .
- Рупперт, Э. Э. Зоология беспозвоночных: Функциональные и эволюционные аспекты : в 4 т. / Э. Э. Рупперт, Р. С. Фокс, Р. Д. Барнс ; под ред. А. А. Добровольского и А. И. Грановича . — М. : Издательский центр «Академия», 2008. — Т. 1. — 496 с. — ISBN 978-5-7695-3493-5 .
- Серавин, Л. Н. / Л. Н. Серавин, А. В. Гудков . — СПб. : ТЕССА, 2005. — 69 с. — (Конспекты по зоологии беспозвоночных ; вып. 1). — ISBN 5-94086-045-1 .
- Schierwater, B. // Key Transitions in Animal Evolution / B. Schierwater, M. Eitel, H.-J. Osigus [et al.] ; Ed. by R. DeSalle and B. Schierwater . — Boca Raton, Florida : CRC Press , 2010. — P. 289—326. — ISBN 978-1-57808-695-5 .
|
Классификация
животных
|
|
|---|---|
| Губки | |
|
|
| Гребневики | |
| Стрекающие | |
| Bilateria | |
- 2021-02-28
- 1