Interested Article - Базидия

- 2020-08-12
- 1

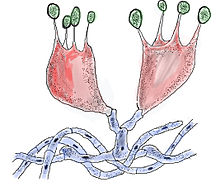
Бази́дия — специализированная структура полового спороношения грибов - базидиомицетов . Базидии представляют собой утолщённые терминальные (концевые) клетки дикариотических гиф или многоклеточные структуры. У экзобазидиальных грибов, не образующих плодовых тел, они формируются в гимении плодовых тел или непосредственно на мицелии ; у головнёвых и ржавчинных грибов базидии возникают из крупных бесполых спор, называемых телиоспорами , телейтоспорами или пробазидиями .
Основное отличие базидий от асков — аналогичных спороносящих структур у аскомицетов — экзогенное (наружное) образование спор ( базидиоспор ), в то время как аски продуцируют споры ( аскоспоры ) эндогенно (внутри). Базидиоспоры образуются на концах узких выростов — стеригм , и после созревания отрываются от них.
Термин базидия ( фр. baside , лат. basidium ) введён в 1837 году Ж.-А. Левейе в работе «Recherches sur l’hymenium des champignons», опубликованной в 8 томе издания «Annales des sciences naturelles»; ранее для базидий и асков использовалось общее наименование тека . Наличие базидий послужило основанием для выделения особого раздела грибов. В 1846 году Ж.-А. Левелье назвал эти грибы Basidiospori , в настоящее время они составляют отдел Базидиомикота ( Basidiomycota ) .
Базидии очень разнообразны по строению, их признаки длительное время использовались в систематике для выделения крупных таксонов базидиомицетов — в XX веке эту группу грибов рассматривали как класс, который разделяли на два или три подкласса в зависимости от наличия одноклеточных холобазидий или разделённых септами фрагмобазидий или гетеробазидий .
В старой литературе «базидиями» также называли некоторые элементы бесполого ( конидиального ) спороношения — конидиеносцы или , в таком смысле термин более не используется .
Строение
В морфологии базидий выделяют три основные части:
- тело , которое может быть одноклеточным или состоять из нескольких клеток — гаплоцитов ;
- стеригмы — узкие выросты, число которых равно числу производимых базидией спор;
- базидиоспоры .
Стеригмы обычно имеют форму узкого конуса, часто расположены на верхушке цилиндрической или булавовидной базидии, но могут образовываться и на боковых поверхностях. У многоклеточных базидий каждый гаплоцит формирует одну стеригму.
В терминах, обозначающих различные части или стадии развития базидии, у разных авторов существовали значительные расхождения. Эта терминология была пересмотрена в 1973 году и в 1988 году Х. Клеменсоном , которые предложили использовать нижеследующие определения (главным образом соответствующие терминологии М. А. Донка [1931, 1954]).
- Пробазидия ( первичная базидиальная клетка , пробазидиальная циста ) — морфологическая часть или стадия развития, в которой происходит кариогамия (слияние двух ядер дикариона). У В. Нойхоффа (1924), (1934), Дж. Мартина (1938) этой структуре соответствует термин гипобазидия . Пробазидия соответствует телиоспоре ржавчинных грибов ( Pucciniales ).
- Метабазидия — морфологическая часть или стадия развития, в которой осуществляется мейотическое деление ядра. У Нойхоффа, Роджерса, Мартина (1938) метабазидии соответствует термин эпибазидия , рекомендуемый «Словарём грибов» для обозначения стеригмы, а согласно Мартину (1957) метабазидия вместе с пробазидией составляют гипобазидию . Для случаев, когда метабазидия включает остаток пробазидии, удалённая от него функционирующая часть может называться париобазидией .
- Стеригма — вырост метабазидии, состоящий из протостеригмы (Донк), или эпибазидии (Мартин, 1957) — узкой или вздутой базальной части и спикулюма , или собственно стеригмы — апикального (концевого) выступа, несущего базидиоспору .
Базидиоспоры обладают билатеральной симметрией и дорсовентрально несимметричны , то есть имеют более выпуклую дорсальную сторону и уплощённую вентральную . Для большинства грибов характерна адаксиальная ориентация спор относительно оси базидии, то есть дорсальными поверхностями они повёрнуты наружу, противоположная абаксиальная ориентация встречается значительно реже .
Формирование и развитие
Известно три типа образования базидий .
- Пряжкой . Пряжки в основании базидий, как и в других частях мицелия, служат для распределения ядер делящегося дикариона между формирующейся пробазидией и её клеткой-ножкой.
- Почкованием . Некоторые виды не имеют пряжек, и базидии возникают из концевых клеток гиф или их ответвлений.
Сходные и близкородственные виды могут отличаться способностью образовывать пряжки и участием пряжек в образовании базидий, такие различия могут проявляться между разными видами одного рода. Например, у пряжки полностью отсутствуют, у имеются только в основаниях базидий, а у пряжки образуются возле каждой септы гиф мицелия, в том числе при базидиях.
- Пробазидиальный . Пробазидия может представлять собой отросток терминальной клетки гифы, не отделённый септой, например, у из класса Pucciniomycetes . Дикарион перемещается в этот отросток и здесь происходит кариогамия, затем диплоидное ядро вместе с клеточной плазмой возвращается в растущую гифу, которая развивается в метабазидию. У других видов пробазидия может формироваться как вздутая толстостенная клетка гифы, которая после кариогамии прорастает метабазидией. В метабазидию перемещается диплоидное ядро и здесь осуществляется мейоз и образование спор, а пробазидия остаётся безъядерной. У ржавчинных и головнёвых грибов пробазидиями служат телиоспоры.
У гименомицетов терминальные клетки гимениальных гиф формируют базидиолы — клетки, сходные с базидиями, но не имеющие стеригм. Базидиолы могут иметь некоторый период покоя, после которого в них осуществляется кариогамия и мейоз, а затем формируются стеригмы и споры. В этом случае пробазидия и метабазидия являются стадиями развития, а не морфологическими частями базидии.
Спорогенез и отделение спор
После мейотического деления диплоидного ядра базидии и происходящего затем у некоторых видов митоза на концах стеригм образуются споровые пузыри с растянутыми стенками, в которые переходят обычно по одному дочернему ядру и часть цитоплазмы, затем формирующаяся спора покрывается плотной оболочкой .
«Типичными» базидиями считаются четырёхспоровые , то есть такие, у которых не происходит митоз, и 4 ядра, образовавшиеся в результате мейоза, формируют споры. Четырёхспоровые базидии, однако, могут образовываться и при осуществлении дополнительного митотического деления дочерних ядер, в этом случае 4 из 8 ядер дегенерируют.
Многоспоровые ( мультиспоровые ) базидии образуются у видов, для которых характерен постмейотический митоз всех или части дочерних ядер, и все ядра после этого формируют споры. Известны 6-споровые (например, у ) и 8-споровые ( и другие) базидии; если митоз происходит два или три раза, возникают 16-споровые и 32-споровые ( ) базидии. Существуют также нетипичные базидии, обладающие неограниченным ростом и образующие неопределённое количество (несколько десятков) спор ( , Mixia ). У некоторых видов базидиоспоры способны к почкованию после созревания, но ещё до отделения их от базидии.
Двуспоровые ( биспоровые ) базидии могут образовываться по одному из трёх известных механизмов.
- Дочерние ядра сразу объединяются в дикарионы и образуют две дикариотические споры, поэтому стадия монокариотического (первичного) мицелия у таких грибов отсутствует. Такой механизм характерен для шампиньона двуспорового ( Agaricus bisporus ). Шампиньон двуспоровый и другие виды со сходным жизненным циклом называют псевдогомоталлическими , их мицелий, проросший из единственной споры ( моноспоровый изолят ), способен к образованию плодовых тел без осуществления плазмогамии с мицелием-партнёром .
- Два из четырёх гаплоидных ядер дегенерируют и не формируют споры. Этот механизм наблюдается у , .
- Четыре ядра осуществляют митоз, но из полученных 8 ядер только два образуют монокариотические споры (у клавулины гребенчатой ( Clavulina cristata ), вороночника рожковидного ( Craterellus cornucopioides )) или 4 ядра образуют дикариотические споры ( , ), а остальные ядра дегенерируют.
У многих видов грибов наряду с чётноспоровыми базидиями в гимении встречаются аберрантные базидии с нечётным числом спор — 3 (у белого гриба ( Boletus edulis )), 5 ( Навозник белоснежный ( Coprinopsis nivea )) или 7 ( Лисичка обыкновенная ( Cantharellus cibarius )). Предполагают, что такие аберрации вызываются дегенерацией одного из ядер в результате проявления в гаплоидном состоянии летальных аллелей .
Отделение базидиоспор чаще всего происходит активно, они отстреливаются от стеригмы обычно на расстояние 0,1—0,3 миллиметра, то есть достаточное для того, чтобы попасть в просвет трубочки или между пластинками гименофора, затем спора падает и уносится потоком воздуха. Для гастеромицетов, очевидно, активное отделение спор не имеет смысла, и у них механизм отстреливания редуцирован. Базидиоспоры, отделяющиеся активным способом, называют баллистоспорами , а отделяющиеся пассивно — статисмоспорами . Вместе со спорой может отделяться и часть стеригмы, которая служит для увеличения летучести споры.
В основании споры при её созревании образуется гликопротеиновая пробка ( англ. plug ), с двух сторон ограниченная линзовидными перегородками, сформированными клеточной стенкой — крышечкой ( англ. lid ) и шапочкой ( англ. calotte ). Пробка набухает от влаги, содержащейся в воздухе, и её содержимое выделяется через отверстие, называемое лат. punctum lacrimans (букв. плакучая точка ). На поверхности в основании споры образуется вязкая апикулярная капля , или капля Буллера , вначале покрытая растягивающимся внешним слоем клеточной стенки стеригмы. Одновременно поверхность споры покрывается тонким слоем конденсирующейся из воздуха влаги, или отдельными капельками. В момент, когда капля Буллера сливается с поверхностной водяной плёнкой, происходит резкое смещение центра масс , что и приводит к отрыву и отстреливанию баллистоспоры. Спора приобретает начальную скорость 30—60 см/сек и преодолевает расстояние до центра просвета гименофора (или несколько меньшее) за 2—3 миллисекунды .
Классификация
Базидии классифицируют в зависимости от положения их по отношению к генеративной гифе, особенностей онтогенеза , наличия или отсутствия внутри базидии (другими словами — разделена ли базидия на гаплоциты) и от ориентации веретён деления ядер при мейозе по отношению к продольной оси базидии.
Принципы классификации
По положению относительно оси генеративной гифы выделяют три типа базидий.
- Апикальные базидии образуются из терминальной клетки гифы и расположены параллельно её оси.
- Плевробазидии образуются из боковых отростков и расположены перпендикулярно оси генеративной гифы, которая продолжает расти и может образовывать новые отростки с базидиями.
- Подобазидии образуются из бокового отростка, повёрнутого перпендикулярно к оси генеративной гифы, которая после формирования одной базидии прекращает свой рост .
В зависимости от дальнейшего развития базидиолы базидии делят на два типа.
- Гомобазидии образуются непосредственно из базидиолы после осуществления в ней кариогамии и мейоза. Тело гомобазидии фактически представляет собой созревшую базидиолу.
- Гетеробазидии образуются путём прорастания базидиолы (пробазидии) после кариогамии, а мейоз осуществляется в новой сформированной структуре — метабазидии. Метабазидия может состоять из 2 или 4 отдельных выростов — протостеригм, каждая из которых формирует одну стеригму (спикулюм). Тело гетеробазидии, таким образом, состоит из двух частей — пробазидии и метабазидии .
В зависимости от наличия или отсутствия септ также выделяют два типа.
- Холобазидии , или голобазидии — одноклеточные.
- Фрагмобазидии разделены септами, обычно на четыре гаплоцита.
Считается, что гетеробазидии являются эволюционно более ранним типом спороношений, чем голобазидии. Септы в базидии не выполняют каких-либо важных функций, поэтому в ходе эволюции они постепенно редуцируются. Известны переходные базидии с частичными перегородками .
Ещё на два типа базидии разделяют в зависимости от расположения ядер после второго деления мейоза.
- У стихических , или стихобазидий веретено первого деления мейоза расположено в плоскости, параллельной продольной оси метабазидии. Веретена второго мейотического деления могут иметь различное расположение, но все дочерние ядра никогда не бывают в верхней части базидии.
- У хиастических , или хиастобазидий оба этапа мейоза происходят в верхней части и на обоих этапах веретена деления перпендикулярны продольной оси метабазидии .
В результате у стихических фрагмобазидий септы разделяют метабазидию в поперечном направлении, а у хиастических — в продольном. У некоторых грибов ( Exobasidium , Tilletia ) хиастические базидии могут после мейоза разделяться поперечными септами, но в таком случае в спорообразовании принимает участие только верхняя клетка. Такие базидии относят к холо -типу. Стихобазидии считаются архаичными структурами. На ранних стадиях развития они сходны с асками и происходят, вероятно, от спороношений, которые были свойственны общим предкам аскомицетов и базидиомицетов. Споры у стихобазидий находятся в неравных условиях — верхним оказывается проще покинуть плодовое тело, чем нижним. Поэтому такие базидии могут иметь изогнутую Г-образную форму, но полностью преодолевается этот нежелательный эффект только при переходе к хиастобазидям .
Основные типы
Объединяя указанные выше принципы классификации, выделяют семь основных типов базидий.
- Agaricus -тип — хиастические гомо-холобазидии;
- -тип — хиастические гетеро-холобазидии;
- -тип — хиастические гетеро-холобазидии, образующиеся из телиоспор;
- Tremella -тип — хиастические гетеро-фрагмобазидии;
- Auricularia -тип — стихические гомо-фрагмобазидии;
- Septobasidium -тип — стихические гетеро-фрагмобазидии;
- -тип — стихические гетеро-фрагмобазидии, образующиеся из телиоспор.
| Холобазидии | Фрагмобазидии | ||
| Хиастобазидии | Стихобазидии | ||
| Гомобазидии | Agaricus -тип | — | Auricularia -тип |
| Гетеробазидии | Tulasnella -тип | Tremella -тип | Septobasidium -тип |
| Гетеробазидии с телиоспорами | Tilletia -тип | — | Ustilago -тип |
Внутри этих типов существуют многочисленные вариации, отличающиеся количеством спор, расположением и ориентацией их на базидии, способом отделения спор, положением базидии относительно генеративной гифы, формой тела базидии и стеригм, наличием и расположением септ и др.
Agaricus -тип
Этот тип характерен для классов Агарикомицеты ( Agaricomycetes ) и ( ), иногда встречаются у пукциниомицетов ( Pucciniomycetes ), тремелломицетов ( Tremellomycetes ).
Типичные четырёхспоровые базидии Agaricus -типа имеют мешковидную форму с округлым сечением в средней части и почти четырёхугольным сечением верхней части (апекса). Их верхушка представляет собой «площадку», в углах которой находятся 4 стеригмы, несущие споры в адаксиальной ориентации.
Двуспоровые базидии встречаются у многих шляпочных грибов и у родов , , относящихся к экзобазидиомицетам.
Мультиспоровые характерны, например, для рода и некоторых других агарикомицетов, а также для из экзобазидиомицетов.
Плевробазидии Agaricus -типа встречаются у родов Botryobasidium , и , относящегося к тремелломицетам. У Botryobasidium они шестиспоровые, а у Erytrobasidium двуспоровые.
Базидии с абаксиальной ориентацией спор характерны для представителей порядка экзобазидиальных ( Exobasidiales ) — , , . У этих грибов могут также возникать в теле базидий вторичные поперечные септы, споры у Muribasidiospora также септированы ( муральные ).
Статисмоспоровые базидии характерны для гастеромицетов , иногда такой тип встречается у пукциниомицетов ( , ). У рода Тулостома ( Tulostoma ) стеригмы располагаются не апикально, а хаотически по всему телу базидии .
Tulasnella -тип
Такие базидии встречаются у класса Agaricomycetes , характерны для Tremellomycetes , Dacrymycetes .
Для типичных базидий этого типа характерны округлая (эллипсовидная) пробазидия и протостеригмы широкой веретеновидной (род ) или более узкой и удлинённой шиловидной ( ) формы, расположенные на апикальной площадке. В последнем случае такие базидии трудно визуально отличить от Agaricus -типа. Всё же они отличаются бо́льшей длиной стеригм (являющихся здесь частями метабазидии) и утолщённой клеточной стенкой пробазидии.
Двуспоровые базидии иногда рассматриваются как самостоятельный -тип , характерный для класса Dacrymycetes . Их тело состоит из вытянутой узкой пробазидии и двух протостеригм, приблизительно равных по размерам пробазидии, в целом тело имеет вытянутую вильчатую форму. Стенки пробазидии у этой вариации не утолщённые.
Вариация с полной метабазидией , как и предыдущая, может рассматриваться как отдельный -тип , он характерен для порядка . Пробазидия у этого типа морфологически сходна с телиоспорой, но не способна к распространению. Она шаровидной формы с толстой оболочкой и имеет самостоятельное наименование склеробазидия . Прорастает цилиндрической метабазидией, несущей на апикальной площадке 4—6 спор, причём споры обычно ориентированы горизонтально и расположены по кругу, в целом эта структура напоминает венчик цветка.
Булавовидные многоспоровые базидии с полной метабазидией также иногда выделяют в отдельный -тип , кроме рода Filobasidium , встречающийся у семейства дрожалковых ( Tremellaceae ). Пробазидия не имеет утолщённой клеточной стенки, она вытянутой цилиндрической или веретеновидной формы. Метабазидия сферической формы, несущая на коротких стеригмах 8 и более спор. У рода Filobasidiella споры почкуются, не отделяясь от базидии, и формируют длинные цепочки, которые затем образуют дрожжевую анаморфу — криптококк ( Cryptococcus ) .
Tilletia -тип
Базидии Tilletia -типа распространены у класса Устомицеты ( Ustilaginomycetes ) (роды , , ), встречаются у экзобазидиомицетов ( ) и класса . Споры у этих грибов могут иметь удлинённую веретеновидную или серповидную форму, вероятно, способствующую их распространению с потоками воды. Спор на базидии бывает различное число, но обычно больше четырёх. У рода Tilletia базидии, а у и споры разделяются вторичными поперечными септами. У родов Ramphospora , споры на базидии способны почковаться и образуют пучки вторичных споридий , расположенных на дистальном конце (противоположном месту прикрепления к базидии) такой споры.
Базидии Tilletia -типа с многоклеточными телиоспорами встречаются у представителей рода и немногих устомицетов ( ). После мейоза пробазидия делится на четыре ( Entorrhiza ) или две ( Mycosyrinx ) клетки, каждая из которых затем прорастает. Такие базидии подобны базидиям с неполной метабазидией Tulasnella -типа, отличаются от них пропагативной пробазидией .
Tremella -тип
Типичные базидии этого типа имеют овальную пробазидию, разделённую продольными септами на четыре гаплоцита, каждый из которых прорастает одной цилиндрической протостеригмой с одной спорой. Они характерны для родов Эксидия ( Exidia ), Pseudohydnum , Tremella класса Tremellomycetes .
Базидии с неполными септами встречаются у родов , , . Пробазидия у них разделена только в верхней части, септы не доходят до её основания. Эта вариация считается переходным звеном между Tremella - и Tulasnella -типами .
Auricularia -тип
Внутри этого типа существуют разнообразные вариации. Такие базидии характерны для аурикуляриевых ( Auriculariales ), некоторых представителей пукциниомицетов , образующих дрожалковидные плодовые тела, классикуломицетов ( Classiculomycetes ), атрактиелломицетов ( Atractiellomycetes ), агарикостильбомицетов ( Agaricostilbomycetes ) и некоторых дрожалковых ( Tremellales ).
Типичные Auricularia -базидии встречаются у аурикуляриевых и родов пукциниомицетов , . Их пробазидия веретеновидной формы разделена на 4 гаплоцита поперечными септами, обычно образующими цилиндрические протостеригмы; у рода протостеригмы вздутой веретеновидной формы.
Спиралевидные плевробазидии Auricularia -типа известны у рода Phleogena (атрактиелломицеты).
Двуклеточные (двуспоровые) базидии встречаются у представителей атрактиелломицетов, например рода .
Статисмоспоровые базидии с короткими стеригмами и сферическими спорами отмечены у родов , Phleogena .
Мультиспоровые базидии узкой цилиндрической формы, каждый гаплоцит у них образует грозди из нескольких (около 8) спор. Такие базидии внешне похожи на некоторые конидиальные спороношения (конидиеносцы с ботриобластоконидиями). Они встречаются у агарикостильбомицетов.
Базидии с диагональными септами — уникальные структуры, нечто среднее между стихо- и хиастобазидиями. Они утолщённой веретеновидной формы, септы у них расположены под углом, близким к прямому, по отношению друг к другу и на продольном сечении базидии образуют Z-образную фигуру. Встречаются у некоторых представителей дрожалковых — , .
Пропагативные базидии рода Sirobasidium образуются цепочками на генеративной гифе и отрываются до образования спор. В отличие от телиоспор, они не имеют толстостенной оболочки .
Septobasidium -тип
Этот тип отмечен у некоторых родов пукциниомицетов и у класса Cystobasidiomycetes . Считается вероятным, что он представляет собой древнейший план строения базидий.
Типичные Septobasidium -базидии состоят из непропагативной пробазидии овальной формы и веретеновидной метабазидии с четырьмя гаплоцитами. Наблюдается у родов Septobasidium , , (пукциниомицеты) и .
У вариации с плевральной пробазидией метабазидия формируется в основании, а не на верхушке пробазидии, которая имеет вид мешковидного придатка. Такая вариация характерна для рода .
Ustilago -тип
Типичные базидии характерны для родов устомицетов , таких, как , , для некоторых экзобазидиомицетов — , и многих представителей пукциниомицетов — , , , , и других. Сферическая толстостенная телиоспора этих грибов прорастает веретеновидной метабазидией, состоящей из 3—4 гаплоцитов. У многих видов базидиоспоры на базидии способны почковаться, а иногда и копулировать, после чего дают начало сразу дикариотическому мицелию.
Базидии с отделяющимися метабазидиями известны у порядка , род . Они имеют трёхклеточные метабазидии, которые продуцируют почкующиеся споры только после отделения от пробазидий.
Вариация с двуклеточными метабазидиями характерны для рода . Каждый из двух гаплоцитов у этих грибов образует 1—2 базидиоспоры.
Многоклеточные телиоспоры встречаются у многих ржавчинных грибов ( Pucciniomycetes ). У рода ( ) они состоят из двух клеток, у — из трёх, у — из четырёх, у число клеток телиоспоры — 5—8, у более восьми. Такие телиоспоры имеют крупный размер и разнообразны по строению. Каждая их клетка представляет собой отдельную пробазидию, прорастающую четырёхспоровой метабазидией.
Телиоспоры с редуцированной метабазидией известны у нескольких родов пукциниомицетов, например, . Мейоз у них осуществляется внутри оболочки веретеновидной телиоспоры, которая затем формирует гаплоциты, прорастающие односпоровыми стеригмами. Это переходный вариант, имеющий признаки как гетеро-, так и гомобазидии, при этом сохраняется пропагативная функция пробазидии .
Нетипичные базидии
Половые спороношения некоторых базидиомицетов обладают уникальными особенностями, они могут быть сходны по некоторым признакам с асками или вовсе не имеют аналогов. Такие структуры обычно не называют базидиями, а применяют к ним общий для половых спороношений термин мейоспорангии .
Мейоспорангии у рода (класс Pucciniomycetes ) развиваются по Auricularia -типу, но развитие прекращается на стадии образования гаплоцитов. Гаплоциты отделяются друг от друга и целиком выполняют функцию базидиоспор. Сходные мейоспорангии имеет род (порядок дрожалковых ), они отличаются гаплоцитами, формирующимися в тетраэдрах, а не рядами в веретеновидной структуре.
Род из класса Microbotryomycetes образует структуры, сходные с базидиями Ustilago -типа, однако они могут давать как экзогенные, так и эндогенные споры. Такие мейоспорангии могут быть смешанными, то есть часть гаплоцитов у них формируют типичные базидиоспоры, а другие — эндогенные споры.
Мейоспорангии рода из порядка экзобазидальных ( Exobasidiales ) сходны с базидиями Auricularia -типа, но способны к неограниченному росту. Все клетки генеративной гифы, начиная с терминальной, преобразуются в метабазидии, которые затем формируют множество статисмоспор.
Род Mixia из подотдела Pucciniomycotina образует крупные конусовидные структуры, на всей поверхности которых развивается большое число спор .
Морфология базидий и систематика базидиомицетов
В XX веке в систематике базидиомицетов признаки строения базидий использовались для выделения подклассов в классе Basidiomycetes , после повышения базидиомицетов до ранга подотдела (в 1970-х годах) эти подклассы стали классами. Параллельно существовали две близкие системы. Согласно одной из них, предложенной в 1900 году Н. Т. Патуйяром , базидиомицеты разделяли на подклассы Гомобазидиомицеты ( Homobasidiomycetidae ) и Гетеробазидиомицеты ( Heterobasidiomycetidae ); другая система, сторонником которой был , включала подклассы Холобазидиомицеты ( Holobasidiomycetidae ) и Фрагмобазидиомицеты ( Phragmobasidiomycetidae ) (позже — классы Homobasidiomycetes и Heterobasidiomycetes или Holobasidiomycetes и Phragmobasidiomycetes ). В 1971 году был введён новый класс Телиомицеты Teliomycetes , объединивший грибы с базидиями, вырастающими из телиоспор. В 1983 году разработана система, учитывающая не только строение базидий, но и комплекс признаков, в который входит также строение септы, наличие пряжек мицелия, особенности жизненного цикла, тип полового процесса, наличие плодовых тел. Из «старых» классов в системе Крайзеля были сохранены только телиомицеты. С 1990-х годов разрабатывается система, в основу которой положена система Крайзеля, но учитываются также данные молекулярно-филогенетических исследований. В новой систематике класс Teliomycetes уже не используется. Строение базидий, однако, продолжает играть важную роль в комплексе признаков, используемом в современной систематике .
Примечания
- , с. 421.
- , с. 309.
- , с. 392.
- ↑ , p. 79—82 (basidium).
- , с. 192.
- Talbot P. H. B. Towards uniformity in basidial terminology // Transactions of the British Mycological Society. — 1973. — Vol. 61. — P. 497—512. — doi : .
- , p. 665 (sterigma).
- , с. 193.
- , с. 269.
- , с. 274.
- , с. 155.
- , с. 194—195.
- , с. 193—194.
- , с. 274—275.
- ↑ , с. 195.
- , с. 196.
- , с. 272—273.
- , с. 197.
- , с. 197—198.
- , с. 199—200.
- , с. 200—201.
- , с. 201.
- , с. 202.
- , с. 202—203.
- , с. 204.
- , с. 204—205.
- , с. 205—206.
- , с. 404.
Литература
- Kirk P. M., Cannon P. F., Minter D. W., Stalpers J. A. et al. Ainsworth & Bisby's Dictionary of the Fungi. — CAB International, 2008. — 771 p. — ISBN 978-0-85199-826-8 .
- Мюллер Э., Лёффлер В. / пер. с нем. К. Л. Тарасова. — М. : "Мир", 1995. — С. —274. — ISBN 5-03-002999-0 .
- Кутафьева Н. П. Морфология грибов: Учебное пособие. — Красноярск: изд-во Красноярского ун-та, 1999. — ISBN 5-7638-0161-X .
- Леонтьєв Д. В., Акулов О. Ю. Загальна мікологія: Підручник для вищих навчальних закладів. — Харків: «Основа», 2007. — С. 192—206. — ISBN 978-966-495-040-1 . (укр.) (Общая микология: Учебник для высших учебных заведений)
- Курсанов Л. И. Микология. — М. : Учпедгиз, 1940.
- Курс низших растений / под общей редакцией М. В. Горленко . — М. : "Высшая школа", 1981.
- Ботаника: Курс альгологии и микологии: Учебник / под ред. Ю. Т. Дьякова. — М. : изд-во МГУ, 2007. — ISBN 978-5-211-05336-6 .
|
|
|
|---|
- 2020-08-12
- 1