Interested Article - Молекулярная эволюция

- 2020-08-12
- 2
Молекулярная эволюция ( англ. molecular evolution ) — наука, изучающая процесс изменения последовательностей мономеров в биополимерных молекулах в живых организмах, а именно ДНК , РНК и белков . Молекулярная эволюция опирается на принципы эволюционной биологии , молекулярной биологии и популяционной генетики . Задача молекулярной эволюции состоит в объяснении закономерностей таких изменений. Молекулярная эволюция занимается механизмами накопления изменений молекулами, и механизмами закрепления этих изменений в популяциях, а также проблемами видообразования .
Связь с областями науки
Молекулярная эволюция имеет тесную связь с областями науки:
- палеонтология (датировка эволюционных событий)
- генетика (принципы организации и передачи наследственной информации)
- молекулярная биология (строение генетических макромолекул)
- биофизика (механизмы функционирования генетических макромолекул)
- математика (построение моделей эволюции)
- эволюция (общие эволюционные закономерности)
- информатика (обработка и анализ данных)
- биохимия (химическая природа и превращение генетических макромолекул) .
Объекты исследования
- Последовательности нуклеиновых кислот (ДНК и РНК) как носителей генетической информации
- Последовательности белков
- Структура белков
- Геномы организмов
Методы анализа
Выделяют следующие методы молекулярной биологии :
- гибридизация ДНК
- иммунологические тесты (используемые для определения групп крови)
- определение последовательности аминокислот (для гемоглобина , миоглобина , цитохрома с и т. п.)
- секвенирование ДНК
- электрофорез (для различных ферментов )
- биохимическая систематика (использующая разного рода вторичные метаболиты растений, животных и иных организмов)
Основные задачи
- Выявление закономерностей эволюции генетических макромолекул
- Реконструкция эволюционной истории генов и организмов
Механизмы эволюции генома
Основным источником накопления изменений генетического материала являются геномные изменения. Главные механизмы эволюции генома следующие:
Мутации
Основная статья: Мутации
Мутация — это стойкое изменение генома. Мутации появляются в результате ошибок
репликации
, воздействия
радиации
, мутагенных химических соединений, таких как
колхицин
и
эпоксибензантрацен
, или иных видов биологического стресса, воздействия
транспозонов
или
вирусов
. Мутации подразделяются на геномные, генные и хромосомные. Генные мутации — это изменения происходящие внутри одного отдельного
гена
. Хромосомные мутации затрагивают участки одной
хромосомы
. Геномные мутации касаются целых хромосом
. Большинство мутаций происходят в виде однонуклеотидных полиморфизмов, которые представляют собой замену одного
нуклеотида
, что приводит к точечной мутации. Другие типы мутаций приводят к модификации больших по размеру фрагментов ДНК, и могут вызывать
дупликации
,
делеции
,
инсерции
,
инверсии
или
транслокации
.
Для большинства организмов характерно соответствие между типом мутаций и ГЦ-составом.
Транзиции
(мутации, в которых
пуриновое основание
меняется другим пуриновым, а
пиримидиновое
-- другим пиримидиновым) более распространены чем трансверсии (мутации при которых пурин заменяется а пиримидин и наоборот)
. Также малохарактерны мутации, при которых меняется
аминокислота
в итоговом белке-продукте
трансляции
. Мутагенный процесс является стохастическим. Мутации появляются случайно. Вероятность появления мутации в одной нуклеотидной сайте очень мала и составляет для разных организмов около
на сайт для одного поколения.
В то же время для некоторых
вирусов
характерна более высокая частота мутаций, вероятность появления мутации достигает
. Среди всех мутаций можно выделить нейтральные и позитивные. которые не элиминируются из
популяции
, хотя некоторые из них могут быть элиминированы с помощью
дрейфа генов
. Остальные мутации являются негативными и в ходе естественного отбора элиминируются из популяции.
Поскольку мутации являются чрезвычайно редким явлением, они в аккумулируются очень медленно. В то время как количество мутаций, которые появляются в одном поколении может варьировать, в течение длинных временных промежутков мутации аккумулируются на регулярной основе. Используя среднее количество мутаций за одно поколение и разницу между двумя нуклеотидными последовательностями, можно оценить время эволюции с помощью
молекулярных часов
.


Рекомбинация
Основная статья: Рекомбинация
Рекомбинация — это процесс, который приводит обменам нуклеотидных последовательностей между хромосомами или регионами хромосом. Рекомбинация противодействует физической связи между соседними генами. В результате независимое наследование генов приводит к более эффективному отбору, что означает, что в участках с более высоким уровнем рекомбинации будет меньше вредных мутаций, больше эволюционно предпочтительных вариантов, и меньше ошибок при репликации и репарации. Рекомбинация также может генерировать определённые типы мутаций, если хромосомы не выравнены .
Мобильные элементы
Основная статья: Мобильные элементы
Мобильные элементы составляют значительную часть генома и представлены транспозонами , ретротранспазонами и повторяющимися элементами. В норме активность передвижения мобильных элементов подавляется при помощи piРНК , метилрования ДНК . Однако при стрессовых ситуациях и изменении эпигенетического ландшфта клетки, мобильные элементы начинают перемещаться или копироваться в новые участки генома. Транспозоны могут влиять на структуру и функции генов и на эпигенетический контроль их экспрессии . В частности, встраивание транспозона внутрь гена может привести к сдвигу рамки считывания и его поломке. Такие нерабочие гены выходят из под пресса естественного отбора и быстро накапливают мутации, становясь псевдогенами . Часто активность ретротранспозонов приводит к дупликации генов .
Перемешивание экзонов и альтернативный сплайсинг
Основная статья: Альтернативный сплайсинг
Некодирующие последовательности, которые транскрибируются вместе с геном, но в дальнейшем удаляются из первичного транскрипта, называются интронами . Интроны встречаются у всех эукариот и за редким исключением отсутствуют у прокариот . Во времы процессинга РНК происходит сплайсинг , в результате которого интроны вырезаются, а оставшиеся кодирующие участки ( экзоны ) сшиваются в одну молекулу. При альтернативном сплайсинге не все экзоны попадают в зрелую мРНК, и с одного транскрипта может быть получено несколько различных мРНК ,каждая из которых будет соответствовать собственному белку. Альтернативный сплайсинг считается очень важным шагом к повышению эволюционной гибкости эукариот, так как в случае мутаций, приводящих к образованию новых экзонов, появляется новая изоформа гена без потери изначального белка. Немаловажную роль в создании новых генных структур вкладывает .
Псевдогены
Псевдогены представляют собой ранее функционирующие гены, которые по некоторым причинам перестали экспрессироваться. Прекращение экспрессии может быть связано с мутациями в промоторной зоне , с исчезновением старт-кодона , сдвигом рамки считывания или частичной делецией гена. Псевдогенизация гена обычно происходит во время его дупликации. Иногда дуликация не затрагивает промоторную зону гена, поэтому копия с самого начала не может экспрессироваться. При полной дупликации гена одна из копий также выходит из-под эволюционного пресса и может стать псевдогеном .
Горизонтальный перенос генов
Основная статья: Горизонтальный перенос генов
Горизонтальный перенос генов заключается в передачи генетический материал организму-непотомку. Такой эволюционный механизм широко распространён среди прокариот, однако встречается и у эукариотических организмов .
Для прокариот характерно наличие плазмид — небольших кольцевых ДНК, способных к автономной от клетки репликации . Размер плазмид колеблется от 1 до 600 тысяч п. о. Обмен плазмидами является важным механизмом горизонтального переноса генов у прокариот и может происходить двумя способами. Во-первых, плазмида может быть захвачена бактерей из окружающей среды без непосредственного контакта с первичным носителем плазмиды. Такой захват называется трансформацией . Трансформация широко используются в молекулярной биологии, в частности для клонирования целевых генов в прокариотическую клетку. Во-вторых, плазмида может передаваться при помощи конъюгации , при которой бактерии образуют пару и через специальный пиль одна из бактерий передаёт копию своей F-плазмиды .
Помимо передачи плазмид, горизонтальный перенос осуществляется у бактерий трансдукцией . Процесс заключается в переносе участков ДНК между клетками посредством бактериофагов .
Удвоение генома
Основная статья: Полиплоидия
Удвоение генома обычно связано с нарушениями расхождения хромосом во время мейоза . Получившийся в результате удвоения генома организм называется полиплоидом. Полиплоидов можно разделить на две группы:
- Автополиплоиды образуются в результате передачи двух одинаковых наборов хромосом.
- Аллополиплоиды образуются при слиянии геномов двух неродственных видов, обычно разных видов внутри одного рода. В результате геном аллополиплоида можно разделить на два субгенома, а каждый ген диплоидного набора представлен парой гомеологичных генов аллоплоида. В гомеологичной паре каждый ген соответствует одному из родителей.
Первые несколько поколений после удвоения генома происходят масштабные перестановки в геноме полиплоида. Так как каждый ген имеет две копии, некоторые хромосомы могут выпасть из генома с потерей лишь одного варианта генов . Такое явление называется анеуплоидией . Также активизируются перемещения мобильных элементов. Со временем геном полиплоидов приходит в стабильное состояние, и в ходе дальнейшей эволюции пары гомеологичных генов расслаиваются. Чаще всего один ген из пары становится псевдогеном, а второй продолжает выполнять начальную функцию. Иногда пары расходятся в функциях и начинают работать параллельно.
Полиплоидия является мощным механизмом симпатрического видообразования и широко распространена среди грибов и растений . Единичные случаи полиплоидии встречаются у животных.
Полиплоидия сыграла значительную роль в формировании многих современных цветковых . На данный момент считается, что все цветковые растения прошли как минимум через два цикла полиплоидизации .
Размер генома
На размер генома организма, помимо числа генов, влияет число повторяющихся фрагментов ДНК. Суммарный размер генома часто не соотносится со «сложностью» организма. Это связано с тем, что доля транспозонов в геноме может быть очень большой. Кроме того, число генов также не всегда связано с числом стадий развития и тканей организма.
На данный момент нет доказательств того, что размер генома находится под жёстким отбором у многоклеточных эукариот. Размер генома, вне зависимости от числа генов в нём, слабо коррелирует с большинством физиологических параметров. Значительная часть эукариотов, в том числе и млекопитающие , несут огромное количество повторяющихся элементов.
Один из редких случаев действия естественного отбора на уменьшение размера генома отмечен у птиц . В отличие от млекопитающих, эритроциты птиц имеют ядра , которые при значительных размерах замедляют транспорт кислорода . Для поддержания высокого уровня метаболизма , необходимого для полёта, геном птиц уменьшился. Есть косвенные доказательства того, что такие же маленькие геномы были и у всех прочих теропод , что согласуется с эндотермией и высоким уровнем метаболизма динозавров .
Происхождение генов de novo
Новые гены возникают несколькими генетическими механизмами, к которым относятся удвоение генов, ретротранспозиция, образование химерных генов, использование некодирующих последовательностей.
Удвоение генов вначале ведёт к избыточности генома. Позднее изначально одинаковые копии могут разойтись и выполнять уже различные функции. В дополнение к удвоению гена, может происходить удвоение лишь одного домена белка, что приводит к образованию белка с иной доменной архитектурой .
De novo появление гена может также происходить из ранее некодирующей ДНК . К примеру, было зафиксировано 5 случаев возникновения новых генов из некодирующих последовательностях у D. melanogaster . De novo гены также были обнаружены у дрожжей , риса и у человека. Мутация в стоп кодоне может привести к более поздней остановке трансляции транскрипта и к расширению гена за счёт некодирующей последовательности.
De novo эволюция генов может быть воспроизведена в лаборатории. К примеру, удалось вывести новый ген, энтеробактин эстеразу, которая компенсировала аналогичный ген, удалённый у E. coli. Новый белок не был родственен природному ферменту и был длиной всего лишь 100 а.о. вместо 400 а.о .
Движущие силы эволюции
Существует три гипотезы. объясняющие молекулярную эволюцию.
Селекционная гипотеза
Согласно селекционной гипотезе именно селекция является движущей силой молекулярной эволюции. В то время как известно, что большинство мутаций — нейтральные, селекционисты связывают изменения в частотах нейтральных аллелей с неравновесным сцеплением генов, которые находятся в процессе отбора, а не со случайным генетическим дрейфом. Разница в использовании кодонов объясняется способностью даже слабого отбора формировать молекулярную эволюцию.
Нейтральная гипотеза
Нейтральные гипотезы подчёркивают важность мутаций, очищающего отбора и случайного генетического дрейфа.
Введение нейтральной теории Кимурой
, которое последовало за открытиями Кинга и Джукса
, привело к ожесточённым дебатам об актуальности неодарвинизма на молекулярном уровне. Нейтральная теория молекулярной эволюции предполагает, что большинство мутаций в ДНК находятся в местах, не важных для жизнедеятельности организма и его приспособленности. Эти нейтральные изменения фиксируются в популяции. Позитивные изменения будут очень редкими, и поэтому не будут в значительной степени способствовать полиморфизму ДНК.
Вредные мутации не вносят большой вклад в разнообразие ДНК, потому что они негативно влияют на приспособленность организма и поэтому быстро удаляются из генофонда.
Эта теория является основой для молекулярных часов.
Судьба нейтральных мутаций определяется генетическим дрейфом и способствует как нуклеотидному полиморфизму, так и фиксированным различиям между видами.
В наиболее строгом смысле нейтральная теория не точна. Небольшие изменения в ДНК очень часто имеют последствия, но иногда эти эффекты слишком малы для того чтобы действовал естественный отбор. Даже синонимичные мутации не обязательно являются нейтральными , потому что разные кодоны присутствуют в разных количествах, что влияет на скорость трансляции. Также существует теория, именуемая почти нейтральной теорией. Эта теория расширила перспективу нейтральной теории, предполагая, что некоторые мутации являются почти нейтральными, что означает, что как случайный дрейф, так и естественный отбор влияют на динамику. Основное различие между нейтральной теорией и почти нейтральной теорией заключается в том, что последняя фокусируется на слабом отборе, а не строго на нейтральном.
Мутационная гипотеза
Мутационная гипотеза обращает внимание прежде всего на случайный дрейф и смещения в паттернах мутаций . был первым, кто предложил современный взгляд на мутации. Он предположил, что изменение ГЦ состава было не результатом положительного отбора, а следствием мутационного давления ГЦ .
Эксперименты в области молекулярной эволюции in vitro
Принципы молекулярной эволюции были открыты, изучены и в дальнейшем проверены с помощью амплификации, вариации и селекции наиболее быстро пролиферирующих и генетически наиболее вариабельных видов вне клетки. После с новаторской работы
Соломона Шпигельмана
1967 года
, описывающей РНК , которая реплицируется с помощью фермента из вируса Qß
, несколько групп (таких как группа Крамерса
и группа Байбрехера, Луса и Ойгена
) изучили мини- и микро-варианты этой РНК в 1970-х и 1980-х годах, которые самореплецируются в за секунды или минуты, что позволяет проследить за сотнями поколений достаточно большого размера (например, размером в 10 ^ 14 последовательностей) за один день экспериментов.
Химико-кинетический анализ подробного механизма репликации
показал, что этот тип системы был первой молекулярно-эволюционной системой, которую можно полностью охарактеризовать на основе физико-химической кинетики, построив первые модели, которые могли бы позволить установить соответствие от генотипа к фенотипу. Такая модель основана на зависящем от нуклеотидной последовательности сворачивании РНК и рефолдинге
.
При условии сохранения функции многокомпонентного фермента Qß химические условия могут значительно варьироваться, чтобы изучить влияние изменения окружающей среды и давления отбора. Эксперименты с квазивидами РНК in vitro включали характеристику порога ошибки для информации в молекулярной эволюции, открытие эволюции de novo, приводящей к разнообразным реплицирующимся видам РНК, и открытие пространственных бегущих волн как идеальных реакторов молекулярной эволюции.
В более поздних экспериментах использовались новые комбинации ферментов для выяснения новых аспектов взаимодействующей молекулярной эволюции, включающей популяционно-зависимую приспособленность, включая работу с искусственно созданной молекулярной моделью "добыча-хищник"
и кооперативными системами множества РНК и ДНК. Для этих исследований были разработаны специальные эволюционные реакторы, начиная с машин с последовательным переносом, проточных реакторов, таких как машины с клеточной статистикой, капиллярные реакторы и микрореакторы, включая реакторы с линейным потоком и гелевые реакторы. Эти исследования сопровождались теоретическими разработками и моделированием, включающими фолдинг РНК и кинетику репликации, которые выяснили важность структуры корреляции между расстоянием в пространстве последовательностей и изменениями приспособленности, включая роль нейтральных сетей и структурных ансамблей в эволюционной оптимизации.
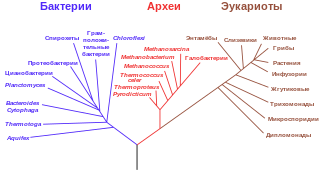
Молекулярная филогенетика
Основная статья: Молекулярная филогенетика
Молекулярная систематика появилась вследствие совмещения традиционной систематики с подходами молекулярной генетики . Молекулярная систематика использует последовательности ДНК, РНК или белков для разрешения вопросов систематики, а именно правильной классификации или таксономии с точки зрения эволюционной биологии.
Молекулярная систематика распространилась из-за доступности техник секвенирования ДНК, которые позволяют определять конкретные последовательности нуклеотидов ДНК или РНК. Секвенирвание всего генома на данный момент всё чаще используется для филогенетических исследований, однако для построения филогении обычно достаточно сравнить всего несколько вариабельных фрагментов длиной 1000 п. о. Такими вариабельными фрагментами часто выступают 16S рРНК у бактерий, и хлоропластные маркёры у растений, митохондриальные фрагменты у животных.
Эволюция белковых последовательностей
Эволюция белков изучается сравнением последовательностей и структур белков из многих организмов отражающим филогенетически удалённые клады. Если последовательности и структуры двух белков схожи, что значает их общее происхождение, то такие белки называются гомологичными . Если гомологичные белки получены из разных видов, то они назывются ортологами. Гомологичные белки, обнаруженные в одном геноме, называются паралогами.
Эволюция белков всегда обусловлена изменениями в ДНК кодирующих белки генов. Но не всегда мутации ДНК отражаются на белковой последовательности, так как часто замена нуклеотида в кодирующем аминокислоту триплете приводит к возникновению сининомичного триплета .
Филогенетические связи белков устанавливаются множественным сравнением последовательностей и построением филогенетических деревьев . Такие филогенетические деревья показывают, что высокое сходство последовательностей отражает эволюционную близость белков.
Эволюция белков описывает изменения во времени формы, функций и устройства белков. Примечательно, что скорость изменения последовательностей неодинаковая у разных белков . Ключевые белки, необходимые для жизнедеятельности организма, обычно более консервативны, так как большинство мутаций в таких генах приводят к значительным нарушениям в работе всего организма и не закрепляются в популяции. В свою чередь, частота мутаций внутри последовательностей одной функциональной группы белков различается между локусами. В ферментах наиболее консервативны остатки активого центра.
См. также
Примечания
- ↑ www.pereplet.ru. Дата обращения: 22 апреля 2020. 9 августа 2018 года.
- (англ.) . postnauka.ru. Дата обращения: 22 апреля 2020. 18 сентября 2020 года.
- . Дата обращения: 28 октября 2010. Архивировано из 20 апреля 2011 года.
- . Дата обращения: 16 июля 2022. 27 мая 2016 года.
- . Дата обращения: 22 апреля 2020. 28 сентября 2020 года.
- . Дата обращения: 19 апреля 2020. 22 октября 2018 года.
- . elementy.ru. Дата обращения: 22 апреля 2020. 1 июля 2019 года.
- M. S. Meselson, C. M. Radding. (англ.) // Proceedings of the National Academy of Sciences. — 1975-01-01. — Vol. 72 , iss. 1 . — P. 358–361 . — ISSN . — doi : . 12 августа 2020 года.
- . www.bioinformatix.ru. Дата обращения: 22 апреля 2020. 18 марта 2022 года.
- Douglas L. Black. (англ.) // Annual Review of Biochemistry. — 2003-06. — Vol. 72 , iss. 1 . — P. 291—336 . — ISSN . — doi : . 12 ноября 2019 года.
- Manyuan Long, Esther Betrán, Kevin Thornton, Wen Wang. (англ.) // Nature Reviews Genetics. — 2003-11. — Vol. 4 , iss. 11 . — P. 865—875 . — ISSN . — doi : . 12 ноября 2019 года.
- . licey.net. Дата обращения: 22 апреля 2020. 14 мая 2018 года.
- А. В. Марков. (рус.) // Доклад в Институте Общей Генетики. — 2008. — 13 ноября. 5 июня 2020 года.
- (англ.) . postnauka.ru. Дата обращения: 22 апреля 2020. 24 сентября 2020 года.
- Стент Г. Молекулярная биология вирусов бактерий. — Москва, 1965(пер. - 1974).
- Justin Ramsey, Douglas W. Schemske. (англ.) // Annual Review of Ecology, Evolution, and Systematics . — Annual Reviews , 2002-11. — Vol. 33 , iss. 1 . — P. 589—639 . — ISSN . — doi : . 25 сентября 2019 года.
- Guillaume Blanc, Kenneth H. Wolfe. (англ.) // The Plant Cell. — 2004-07. — Vol. 16 , iss. 7 . — P. 1667—1678 . — ISSN . — doi : .
- Chris L. Organ, Andrew M. Shedlock, Andrew Meade, Mark Pagel, Scott V. Edwards. (англ.) // Nature. — 2007-03. — Vol. 446 , iss. 7132 . — P. 180—184 . — ISSN . — doi : . 19 декабря 2019 года.
- Aoife McLysaght, Daniele Guerzoni. (англ.) // Philosophical Transactions of the Royal Society B: Biological Sciences. — 2015-09-26. — Vol. 370 , iss. 1678 . — P. 20140332 . — ISSN . — doi : . 3 мая 2020 года.
- M. T. Levine, C. D. Jones, A. D. Kern, H. A. Lindfors, D. J. Begun. (англ.) // Proceedings of the National Academy of Sciences . — National Academy of Sciences , 2006-06-27. — Vol. 103 , iss. 26 . — P. 9935—9939 . — ISSN . — doi : .
- Jing Cai, Ruoping Zhao, Huifeng Jiang, Wen Wang. (англ.) // Genetics. — 2008-05. — Vol. 179 , iss. 1 . — P. 487—496 . — ISSN . — doi : .
- Wenfei Xiao, Hongbo Liu, Yu Li, Xianghua Li, Caiguo Xu. (англ.) // PLOS One / Hany A. El-Shemy. — Public Library of Science , 2009-02-25. — Vol. 4 , iss. 2 . — P. e4603 . — ISSN . — doi : .
- Ann E Donnelly, Grant S Murphy, Katherine M Digianantonio, Michael H Hecht. (англ.) // Nature Chemical Biology. — 2018-03. — Vol. 14 , iss. 3 . — P. 253—255 . — ISSN . — doi : . 25 июля 2019 года.
- Graur, D.; Li, W.-H. Fundamentals of molecular evolution. — , 2000. — ISBN 0-87893-266-6 .
- Casillas, Sònia; Barbadilla, Antonio. Molecular Population Genetics (англ.) // Genetics. — 2017. — Vol. 205 , no. 3 . — P. 1003—1035 . — doi : . — . — PMC .
- Hahn, Matthew W. Toward A Selection Theory Of Molecular Evolution (англ.) // Evolution . — Wiley-VCH , 2008. — February ( vol. 62 , no. 2 ). — P. 255—265 . — doi : . — .
- Hershberg, Ruth; Petrov, Dmitri A. (англ.) // Annual Review of Genetics . — 2008. — December ( vol. 42 , no. 1 ). — P. 287—299 . — doi : . — .
- The Neutral Theory of Molecular Evolution (англ.) . — Cambridge University Press , Cambridge, 1983. — ISBN 0-521-23109-4 .
- Kimura, Motoo. (англ.) // Nature. — 1968. — Vol. 217 , no. 5129 . — P. 624—626 . — doi : . — . — . 3 марта 2016 года.
- King, J.L.; Jukes, T.H. (англ.) // Science . — 1969. — Vol. 164 , no. 3881 . — P. 788—798 . — doi : . — . — . 31 декабря 2019 года.
- ↑ Akashi, H. Weak Selection and Protein Evolution (англ.) // Genetics. — 2012. — Vol. 192 , no. 1 . — P. 15—31 . — doi : . — . — PMC .
- ↑ Fay, J. C., Wu, C. I. (англ.) // Annu. Rev. Genom. Hum. Genet. . — 2003. — Vol. 4 . — P. 213—235 . — doi : . — .
- Nachman M. "Detecting selection at the molecular level" in: Evolutionary Genetics: concepts and case studies (англ.) : journal / C.W. Fox; J.B. Wolf. — 2006. — P. 103—118 .
- The nearly neutral theory expanded the neutralist perspective, suggesting that several mutations are nearly neutral, which means both random drift and natural selection is relevant to their dynamics.
- ↑ Ohta, T. The Nearly Neutral Theory of Molecular Evolution (англ.) // Annual Review of Ecology, Evolution, and Systematics : journal. — Annual Reviews , 1992. — Vol. 23 , no. 1 . — P. 263—286 . — ISSN . — doi : .
- Selectionism and Neutralism in Molecular Evolution (англ.) // . — Oxford University Press , 2005. — Vol. 22 , no. 12 . — P. 2318—2342 . — doi : . — . — PMC .
- Sueoka, N. On the evolution of informational macromolecules // Evolving genes and proteins / Bryson, V.; Vogel, H.J.. — Academic Press, New-York, 1964. — С. 479—496.
- Richard Axel. (англ.) // The American Journal of Cancer. — 1983. 14 сентября 2017 года.
- Shi-Jie Chen. (англ.) // Annual Review of Biophysics. — 2008-06. — Vol. 37 , iss. 1 . — P. 197–214 . — ISSN . — doi : . 22 марта 2019 года.
- Luisiana Cundin. (англ.) // arXiv. — 2010.
- Esteban Domingo, Colin R. Parrish, John J. Holland. . — Elsevier, 2008-06-23. — 573 с. — ISBN 978-0-08-056496-8 .
- Joseph Wright. . — Scientific e-Resources, 2019-11-06. — 310 с. — ISBN 978-1-83947-267-1 .
- Leslie Vega &. . — Scientific e-Resources, 2019-09-13. — 341 с. — ISBN 978-1-83947-450-7 .
- Changbong Hyeon, D. Thirumalai. (англ.) // Biophysical Journal. — 2006-05-15. — Vol. 90 , iss. 10 . — P. 3410–3427 . — ISSN . — doi : .
- Anton Moser. . — Springer Science & Business Media, 2012-12-06. — 480 с. — ISBN 978-1-4613-8748-0 .
- Teruo Fujii, Yannick Rondelez. // ACS Nano. — 2013-01-22. — Т. 7 , вып. 1 . — С. 27–34 . — ISSN . — doi : .
- Shi-Jie Chen. // Annual review of biophysics. — 2008. — Т. 37 . — С. 197–214 . — ISSN . — doi : .
- Justin C. Fay, Chung-I Wu. (англ.) // Annual Review of Genomics and Human Genetics. — 2003-09. — Vol. 4 , iss. 1 . — P. 213—235 . — ISSN . — doi : . 8 марта 2021 года.
Ссылки
- (англ.)
- 2020-08-12
- 2