Interested Article - Гаплогруппа H1 (мтДНК)

- 2021-10-07
- 1
Гаплогруппа H1 — гаплогруппа митохондриальной ДНК человека. Является потомком гаплогруппы H .
Происхождение
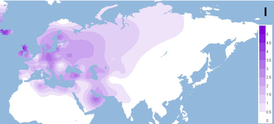
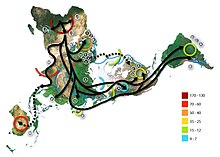
В период последнего ледникового максимума (последнего оледенения) 20-13 тысяч лет назад большинство палеолитических поселений Северной и Центральной Европы вымерло, в связи с чем представители гаплогруппы Н в большей степени выжили лишь на севере современной Испании. Среди современных народов Европы мтДНК гаплогруппа H1 с наибольшей частотой встречается среди басков Испании (27,8 %), португальцев (25,8 %) и жителей Андалусии (24,3 %) . Данные молекулярной генетики говорят о том, что франко-кантабрийский регион был колыбелью большей части населения Европы, по крайней мере, по женской линии (через гаплогруппу H) . Распространение гаплогруппы H1 из франко-кантабрийского региона по всей Европе после последнего ледникового максимума связывают с мадленской экспансией около 13 тысяч лет назад .
Распространение
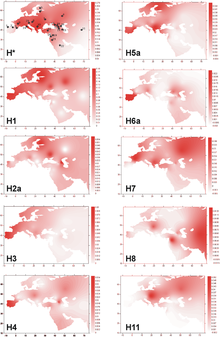
Гаплогруппа H1 составляет значительную долю западноевропейских митохондриальных ДНК, причём пик распространения в Европе приходится на басков Испании (27,8 %), португальцев (25,8 %), жителей Андалусии (24,3 %) и этнографическую группу пасьего в Кантабрии (23,5 %). . В Северной Африке необычно высокая доля гаплогруппы H1 среди туарегов Ливии (61 %). . Также, мтДНК гаплогруппа H1распространена среди других жителей Иберийского полуострова, Северной Африки и Сардинии . Составляет свыше 10 % во многих других регионах Европы (Франция, Британские острова, Альпы, многие регионы Восточной Европы) и не менее 5 % в прочих местах Европы.
Субклады мтДНК гаплогруппы H1
H1a
МтДНК гаплогруппа H1a характеризуется дополнительными мутациями 73G и 16162G. По данным Eva-Liis Loogväli et al. (2004) гаплогруппа H1a встречается среди финнов (5,1 %) словаков (3,4 %), финно-угорских народов Поволжья (2,4 %), французов (1,9 %), эстонцев (1,8 %), восточных славян (1,7 %) и турок (1 %) и не встречается среди народов Ближнего и Среднего Востока и в Азии. Исследуя популяции Ближнего Востока и Кавказа, Roostalu et al. (2007) обнаружили гаплогруппу H1a только у карачаевцев (5,5 %) и турок (0,6 %). Возраст гаплогруппы H1a — 4,5 −9,9 тыс. лет.
H1b
МтДНК гаплогруппа H1b характеризуется дополнительными мутациями 3796G, 16189C и 16356C. При этом, по мнению Mannis van Oven, наличия 16356C уже достаточно, чтобы определять как H1b (даже если нет 3796G).
МтДНК гаплогруппа H1b имеет наибольшее распространение в Восточной Европе (7 % всей гаплогруппы H), на севере Центральной Европы (5 % гаплогруппы H) и на северо-западе Сибири (5 % гаплогруппы H у народа манси ). Пик распространения гаплогруппы H1b в Восточной Европе находится на границе современных Латвии и Литвы, южнее Риги и совпадает с областью расселения балтского племени земгале в Средние Века.
По данным Eva-Liis Loogväli et al. (2004) гаплогруппа H1b встречается среди эстонцев (5,3 %), восточных славян (2,8 %), балканских народов (1,8 %) и словаков (1,7 %) и не встречается среди финно-угорских народов Поволжья, турок, народов Ближнего и Среднего Востока и в Азии.
Несмотря на преобладание гаплогруппы H1b среди народов Восточной Европы, она встречается и на Западе ( испанцы , баски , французы , немцы ), но в значительно меньшей доле. В популяциях Ближнего Востока и Кавказа, Roostalu et al. (2007) обнаружили гаплогруппу H1b у карачаевцев (4,1 %), адыгов (1,1 %), балкарцев (1 %), осетин (0,3 %) и турок (0,3 %). Возраст гаплогруппы H1b — 7,2—14 тыс. лет.
Вероятно, гаплогруппа H1b возникла в южной Франции или в Иберии вскоре после окончания Ледникового периода. Женщины, несущие H1b, путешествовали в восточном направлении с территории современной Франции, прошли к северу от итальянских Альп и вошли в современную Словакию. Оттуда, H1b распространилась на север.
Возможно, на севере Центральной Европы гаплогруппа H1b была связана со свидерской культурой финального палеолита, распространённой в X—IX тыс. до н. э. на территории современных Польши, Литвы и Западной Белоруссии. Носители Свидерской культуры, мигрируя на северо-восток и юго-восток, в VIII тыс. до н. э. могли принести гаплогруппу H1b с севера Центральной Европы на территории современных Эстонии, Южной Финляндии и Центральной России ( бутовская мезолитическая культура Волго-Окского междуречья VIII—VI тыс. до н. э. и кундская мезолитическая культура Восточной Прибалтики VIII—VI тыс. до н. э.).
В митохондриальной гаплогруппе H1b митохондриальная последовательность сына Ефросиньи Мстиславны венгерского короля Белы III имеет самое близкое отношение к человеку из группы KL-VI Карпатского бассейна .
H1f
По данным Eva-Liis Loogväli et al. (2004) гаплогруппа H1f наиболее распространена среди финнов (10,3 %), встречается также у эстонцев (1 %) и восточных славян (0,2 %) и не была выявлена у словаков, французов, турок, финно-угорских народов Поволжья, народов Великобритании, США, Ближнего и Среднего Востока и в Азии. Возраст гаплогруппы H1f около 1000 лет.
H1c
МтДНК гаплогруппа H1c характеризуется дополнительной мутацией 477C. Исследуя популяции Ближнего Востока и Кавказа, Roostalu et al. (2007) обнаружили гаплогруппу H1c только у двух адыгов, что составляет около 2,3 % данной популяции. Возраст гаплогруппы H1c около 9,4 тыс. лет.
H1d
МтДНК гаплогруппа H1d характеризуется дополнительной мутацией 456T, однако данный подкласс был исключён из версии 3 дерева Mannis van Oven. В популяциях Ближнего Востока и Кавказа, Roostalu et al. (2007) обнаружили гаплогруппу H1d только у двух арабов из Кувейта, что составляет около 1,2 % исследованной популяции.
Результаты исследований палеоДНК
Nikitin et al. (Unpub) в пещере Вертеба (Позднее Триполье) в Тернопольской области Украины были обнаружены 2 образца мтДНК гаплогруппы H1 датированные 4300—3100 г.г. до н. э. Lacan et al. (2011), исследуя Treilles (Южная Франция), из 29 образцов выявили 3 мтДНК гаплогруппы H1 датированные 3000 годом до н. э.
У представителя волынцевской культуры из разновременного поселения Куриловка-1 ( Суджанский район Курской области) определили митохондриальную гаплогруппу H1b. Его полный митохондриальный геном оказался полностью идентичен геному венгерского короля Белы III (правил в 1172—1196 гг.), матерью которого была Ефросинья Мстиславна — дочь Великого киевского князя Мстислава и внучка новгородского посадника Дмитра Завидича, и современному геному из Дании (гаплотип 16183C) .
По данным генетиков, у венгерского короля Белы III определена митохондриальная гаплогруппа H1b .
Nilsson et al. в 2010 году исследовали предполагаемые останки католической святой XIV века — Бригитты Шведской . Принадлежность останков святой подвергается некоторому сомнению из-за их радиоуглеродного датирования: возможно, останки относятся к XIII веку, в то время как святая Бригитты жила в XIV веке. МтДНК гаплотип останков (263G, 315.1C, 3010A, 16189C, 16519C) принадлежит гаплогруппе H1b, f, g, k, q , характеризующейся мутацией 16189C.
Бронзовый век
- SI-23 __ EGAN00001390952 / ERS1790729 __ Сидон , Южный Ливан , Ливан __ 1800 BCE (3750 yBP) __ Ж __ H1bc.
Филогенетическое дерево
Приведенное ниже филогенетическое дерево основано на публикации Ван Овена и последующих опубликованных исследованиях.
-
H1
3010A
-
H1a
73G 16162G
- H1a1 6365C 16209
- H1a2 8271T
- H1a3 16051
-
16189C
- H1b 3796G 16189C 16356C
- H1f 4452C 7309C 9066G 16093C
- H1g 8602C
- H1k 5206 15766 16290
- H1q 4859
-
H1c
477C
- H1c1 9150G 16263
- H1c2 8764 12858T
- H2c3 8473
- H1d 456T
-
H1e
5460A
-
H1e1
8512
- H1e1a 14902
- H1e2 15817
-
H1e1
8512
- H1h
- H1i
- H1j
- H1m
- H1n
- H1o 267C 485 6446T 11002 14053G 15844
- H1p 13470G
- H1r 9356
- H1s 8572
-
H1t
9986
- H1t1 14129 14224
- H1t1 5978
- H1u 9923
-
H1v
10314
-
H1v1
4313
- H1v1a 9148
- H1v1b 14560
-
H1v1
4313
- H1w 8966
- H1x 7765 10410A 16037 16256
-
H1a
73G 16162G
Примечания
- ↑ V.Zaporozhchenko (2011), от 30 августа 2011 на Wayback Machine , "Human mtDNA", Data were compiled by V.Zaporozhchenko for the Gentis Company, Build 4, 05.08.2011
- ↑ van Oven, Mannis; Manfred Kayser. (англ.) // : journal. — 2008. — 13 October ( vol. 30 , no. 2 ). — P. E386—E394 . 4 декабря 2012 года.
- ↑ Ottoni et al. 2010, от 13 августа 2011 на Wayback Machine
- Pereira L., Richards M., Goios A., et al. High-resolution mtDNA evidence for the late-glacial resettlement of Europe from an Iberian refugium (англ.) // : journal. — 2005. — January ( vol. 15 , no. 1 ). — P. 19—24 . — doi : . — . — PMC .
- ↑ Дата обращения: 11 сентября 2011. 4 июля 2008 года.
- ↑ Дата обращения: 11 сентября 2011. 19 августа 2009 года.
- ↑ Дата обращения: 12 сентября 2011. 23 сентября 2011 года.
- ↑ от 10 мая 2012 на Wayback Machine , Data from web site of the Gentis Company
- Bea Szeifert et al. от 12 февраля 2022 на Wayback Machine , February 08, 2022
- Дата обращения: 11 сентября 2011. 18 сентября 2011 года.
- Андреева Т. В. и др. , 2023
- Judit Olasz . от 29 сентября 2018 на Wayback Machine , 2018
- Дата обращения: 11 сентября 2011. 11 июня 2012 года.
- Haber, Marc et al. American journal of human genetics vol. 101,2 (2017): 274-282.
- Название данного подкласса — согласно Альваресу-Иглесиасу (Alvarez-Iglesias 2009)
- данный подкласс исключён из версии 3 дерева Ван Овена.
Ссылки
-
- Ian Logan’s
- Provides a phylogenetic tree of global human mtDNA variation.
- National Geographic’s , from National Geographic
- Genebase’s
- Genebase’s
- Genebase’s
- The H1 mtDNA Haplogroup Project
| Митохондриальная Ева | ||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||
| L0 | L3 | |||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||
| M | N | |||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| D | E | G | Q | R | A | S | X | Y | N1 | N2 | ||||||||||||||||
| | | | | | | | | |||||||||||||||||||||||
| C | Z | B | F | R0 | P | I | N1a | W | ||||||||||||||||||
| | | | | | | ||||||||||||||||||||||||
| HV | JT | U | K | |||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| H | V | J | T | Устаревшие кластеры | ||||||||||||||||||||||
- 2021-10-07
- 1