Денитрификация
- 1 year ago
- 0
- 0

Анаэробное окисление метана — процесс окисления метана до углекислого газа , производимый ( англ. ) археями групп , и , близкими к Methanosarcinales при отсутствии в среде молекулярного кислорода . Биохимия и распространённость процесса в природе изучены пока недостаточно.
Уже в первых работах, посвящённых процессу, было показано, что абиотически окисляется лишь 0,7-1,1 % всего окисляемого объёма метана. Также установлено, что ингибиторы метаногенеза и , а также ингибитор сульфатредукции угнетают и анаэробное окисление метана. При этом последний более чувствителен к 2-бромэтансульфоновой кислоте, чем сам метаногенез, из-за чего при небольших концентрациях кислоты она даже оказывает кажущийся стимулирующий эффект на образование метана (на самом деле прекращая его окисление).
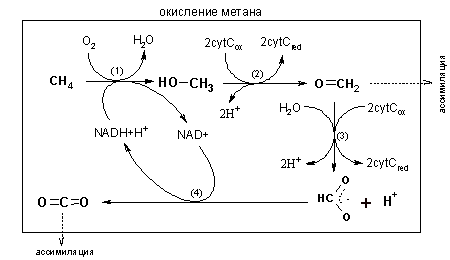
Из этого было сделано заключение о протекании анаэробного окисления метана в два этапа, осуществляемых разными организмами. Первый — окисление метана водой, осуществляемое ферментами метаногенеза, катализирующими обратные реакции. Второй — дальнейшее окисление продуктов первого этапа, с использованием сульфидов в качестве акцептора электрона.
Ингибитором анаэробного окисления метана также является кислород: в образцах донных осадков , где отсутствуют аэробные метанотрофы , при проветривании прекращается всякое окисление метана. Лишь с исчерпанием кислорода в среде оно начинает восстанавливаться.
Благодаря тому что в холодных сипах метан обеднён 13 C , вещества, синтезированные из него, также обеднены этим изотопом. Были изучены бедные 13 C липиды и нуклеиновые кислоты , выделенные из донных отложений, и проведены исследования в области геносистематики потребляющих метан в анаэробных условиях организмов. Было показано, что среди них действительно есть родственные метаногенам порядка Methanosarcinales археи, отнесённые к группам и , и эубактерии-сульфатредукторы . Впоследствии с использованием метода FISH удалось увидеть клетки этих организмов . Они образовывали компактные конгломераты, состоящие в среднем из 100 архей и 200 эубактерий и имеющие диаметр около 3 мкм.
Помимо тесного родства архей ANME метаногенам, доводом в пользу проведения ими обратного метаногенеза стало обнаружение у них генов ( mcrA ) и ещё ряда ферментов образования метана .
На каких продуктах реакции обратного метаногенеза завершаются неизвестно. Видимо часть их превращается в ацетил-КоА и ассимилируется организмом, о чём свидетельствует включение изотопа 13 C в состав микробных биомаркеров. Другая часть поступает во внешнюю среду и усваивается сульфатредукторами, используясь ими затем также в процессах и анаболизма , и катаболизма . Какое соединение выступает в роли межвидового интермедиата неизвестно. На его роль предлагались молекулярный водород , уксусная кислота , метанол , муравьиная кислота , НАД и ФАД . Ни для одного из этих веществ пока не было получено экспериментальных доказательств участия в процессе. Кроме того, термодинамические расчёты свидетельствуют, что водород, ацетат и метанол межвидовыми интермедиатами являться не могут .
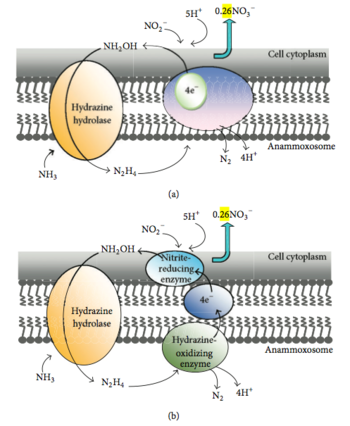
Показана возможность анаэробного окисления метана с одновременным восстановлением нитрата до аммония археями из группы ANME-2d. На данный момент изучение таких микроорганизмов возможно при культивировании в ферментерах. Как правило, их не удается выделить в чистые культуры. Наибольшая концентрация клеток архей в таких накопительных культурах достигает около 80 %. Одним из наиболее изученных представителей является Ca. Methanoperedens nitroreducens. Именно для этого микроорганизма было показано анаэробное окисление метана, связанное с денитрификацией. Кроме того, в последние годы приводится все больше доказательств анаэробного окисления метана, связанного с восстановлением соединений металлов: Fe 3+ , Mn 4+ , Cr 6+ .
Показана возможность протекания анаэробного окисления метана с участием соединений азота . Процесс в этом случае осуществляла консорция клеток ANME-2 и денитрификаторов , причём последние предпочитали нитрит нитратам . При удалении нитрита через 10-20 часов окисление метана возобновлялся уже с участием нитратов. Прямые наблюдения с помощью техники FISH показали, что соотношение в консорции бактерий и архей составило 8:1. Авторы считают, что это свидетельствует о более высоком энергетическом выходе по сравнению с сульфатами.
Для Fe 3+ в нескольких работах утверждается неучастие в анаэробном окислении метана , хотя в других показан его стимулирующий эффект на поглощение метана в анаэробных условиях .
Существует гипотеза наличия одного организма, проводящего процесс от начала до конца по совершенно особому механизму. В её пользу говорят данные, что в некоторых субстратах ни один из ингибиторов метаногенов и сульфатредукторов не подавляет анаэробного окисления метана .
Были обнаружены клетки ANME-2, а затем ANME-1 и ANME-3, физически не связанные с сульфатредукторами, свободноживущие, образующие микроколонии, либо конгломераты клеток с эубактериями, не являющимися сульфатредукторами . Эти организмы теоретически могли бы проводить окисление метана самостоятельно, без участия эубактериального партнёра .
Было обнаружено окисление метана чистыми культурами при анаэробном росте. Метаногены , строгие анаэробы , в особенности такие как , Methanosarcina barkeri , Methanosarcina acetivorans и , способны при росте на обычных средах для культивирования метаногенов, окислять введённый в газовую фазу меченый CH 4 до CO 2 , иногда до метанола и ацетата ( M. barkeri ) в небольших количествах (до 0,3 % от объёма образованного ими за то же время метана) .
Многие сульфатредукторы могут соокислять небольшое количества метана при анаэробном росте на других субстратах. Из них в данном отношении наиболее интересны , имеющие часть ферментов метаногенеза и родственные ANME-1. Проведённые на них исследования, однако, показали отсутствие окисления даже следовых количеств метана . Следует учитывать, что в некоторых работах по сульфатредукторам окисление метана может представлять собой артефакт , вызванный недостаточной чистотой использованного метана, содержащего угарный газ , который и окислялся бактериями .
В любом случае, метанокисляющая активность метаногенов и сульфатредукторов слишком низка, чтобы объяснить с её помощью те объёмы окисляемого метана, о которых свидетельствовали геохимические данные.
Абсолютное большинство исследований по данной тематике выполнено для донных морских и океанических осадков, однако показано, что процесс протекает и в почве , рубце желудка жвачных , пресноводных водоёмах .
Метан является сильным парниковым газом с потенциалом теплопоглощения в 30 раз больше чем у CO 2 . По разным оценкам от 30 до 90 % метана образуется биогенно, в основном в донных осадках и почвах переувлажнённых ландшафтов. Анаэробное окисление метана препятствует его эмиссии в атмосферу. Его масштабы оцениваются в 6-20 % от общего количества ежегодно выделяющегося из океана в атмосферу метана, и предполагается, что эти цифры будут пересмотрены в сторону увеличения. Оценок объёмов анаэробного окисления метана в наземных экосистемах пока не существует.
Первые геохимические свидетельства протекания окисления метана в анаэробных условиях были получены ещё в 1970-е годы при изучении профильного распределения метана, сульфатов и сероводорода в донных отложениях. Первыми из биологов исследовали процесс Зендер и Брок в 1979 — 1980 годах . Ими была выдвинута гипотеза, что окисление протекает в два этапа, причём первый из них представляет собой обратный метаногенез . Позже гипотеза нашла ряд подтверждений и сейчас рассматривается как общепризнанная.
На рубеже 1990-х и 2000-х годов начался новый этап в изучении анаэробного окисления метана с активным привлечением молекулярных методов. Была установлена таксономическая принадлежность анаэробных метанотрофов, начато изучение их генома и кодируемых им ферментов. В 2006 году получены свидетельства, что окисление метана может быть сопряжено с восстановлением не только сульфатов , но и нитратов . Тем не менее, организмы, способные расти в среде с метаном в роли единственного источника углерода, до сих пор не выделены, неясной остаётся биохимия процесса.