Interested Article - Веретено деления

- 2021-08-06
- 1

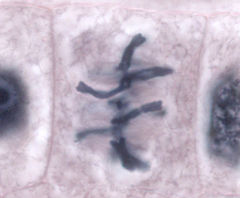
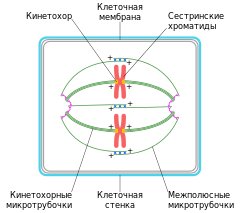
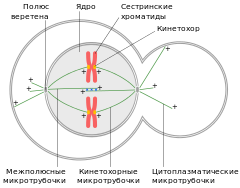
Веретено́ деле́ния — динамичная структура, которая образуется в митозе и мейозе для обеспечения сегрегации (отделения) хромосом и деления клетки. Типичное веретено является биполярным — между двумя полюсами образуется веретенообразная система микротрубочек . Микротрубочки веретена присоединяются к кинетохорам хроматид в области центромер и обеспечивают движение хромосом по направлению к полюсам.
Веретено образуют три основных структурных элемента: микротрубочки, полюса деления и хромосомы. В организации полюсов деления у животных участвуют центросомы , содержащие центриоли . У растений , а также в ооцитах некоторых животных центросомы отсутствуют, и образуется ацентросомальное веретено с широкими полюсами. Важную роль в формировании веретена играют моторные белки , относящиеся к семействам динеинов и кинезинов .
Полноценное веретено деления образуется на стадии прометафазы после разрушения ядерной мембраны , когда цитоплазматические микротрубочки и центросомы (у животных) получают доступ к хромосомам и другим компонентам веретена. Исключение составляет веретено деления почкующихся дрожжей , которое формируется внутри ядра.
Структура
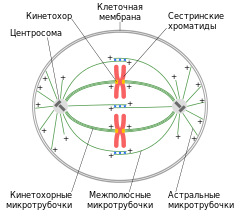
Веретено деления типичной клетки млекопитающих состоит из трёх структурных элементов — центросом , микротрубочек и хромосом , — которые образуют симметричную биполярную структуру. На полюсах веретена располагаются центросомы — небольшие органеллы, функционирующие как центры организации микротрубочек . Каждая центросома состоит из пары центриолей , окруженных множеством разных белков. Между полюсами веретена находятся конденсированные хромосомы, состоящие из пары хроматид , скреплённых в области центромеры . На центромерных участках хромосом находятся кинетохоры — сложные структуры, отвечающие за прикрепление к микротрубочкам веретена .
Веретено деления состоит из двух полуверетён. Полуверетено образуется из поляризованных микротрубочек. Отрицательные минус-концы микротрубочек собираются на полюсах веретена вокруг центросом. Плюс-концы микротрубочек отдаляются от двух полюсов и пересекаются в средней экваториальной части веретена. У большинства позвоночных полуверетено состоит из 600—750 микротрубочек, 30—40 % которых заканчиваются на кинетохорах. Микротрубочки, которые соединяют полюса веретена с кинетохорами хромосом, называются кинетохорными . Причём каждый кинетохор при образовании веретена связывается с множеством микротрубочек и образует кинетохорный пучок. Микротрубочки, которые располагаются между полюсами и не присоединяются к кинетохорам, называются межполюсными . Часть микротрубочек веретена образует вокруг каждого полюса радиальные структуры, называемые звёздами или астерами. Такие микротрубочки называются астральными .
У растений, а также в ооцитах некоторых животных центросомы отсутствуют, и образуется ацентросомальное веретено с широкими полюсами . Также на полюсах ацентросомального веретена отсутствуют астральные микротрубочки. В остальном структура веретена растительной клетки соответствует структуре веретена животной клетки.
Сборка веретена деления
Начало сборки веретена в профазе
Сборка веретена деления начинается в профазе. Однако на данном этапе образование полноценного веретена невозможно по причине изоляции хромосом, а также важных моторных, регуляторных и стабилизирующих белков внутри ядра.
У растений, по причине отсутствия центросом, роль центра организации микротрубочек в профазе выполняет ядерная оболочка. Микротрубочки собираются вблизи поверхности ядра и к окончанию профазы ориентируются вдоль оси будущего веретена деления, образуя так называемое профазное веретено .
В животных клетках центром организации микротрубочек является центросома. Поэтому образование веретена деления начинается с разделения и расхождения пары центросом во время профазы. Расхождение центросом в профазе обеспечивают моторные белки динеины . Они закрепляются на внутренней стороне клеточной мембраны и на внешней поверхности ядра. Закреплённые в мембране динеины присоединяются к астральным микротрубочкам и движутся в направлении минус-конца микротрубочки. За счёт этого центросомы перемещаются к противоположным участкам клеточной мембраны и расходятся дальше друг от друга .
Сборка веретена в прометафазе

1 . Нуклеация микротрубочек вблизи хромосом.
2 . Соединение и сортировка антипараллельных микротрубочек при участии кинезина-5.
3 . Кинезины-4 и -10, закреплённые на плечах хромосом, также участвуют в сортировке и отдаляют минус-концы микротрубочек.
4 . Минус-концы микротрубочек скрепляются между собой и образуют полюса деления при участии динеина и кинезина-14.Сборка веретена зависит от двух ключевых процессов. Во-первых, от формирования биполярного скопления микротрубочек вокруг хромосом. Во-вторых, от прикрепления хромосом к микротрубочкам от противоположных полюсов деления . Присоединение сестринских хроматид к микротрубочкам является неотъемлемой частью процесса сборки веретена. Однако, хромосомы и многие моторные и другие белки, участвующие в формировании полноценного веретена деления изолированы внутри клеточного ядра. А микротрубочки и центросомы (у животных) находятся в цитоплазме. Таким образом, сборка веретена зависит от разрушения ядерной оболочки в прометафазе .
Исключение составляет веретено деления почкующихся дрожжей, которое формируется внутри ядра .
Самоорганизация веретена
У всех эукариот сборка биполярного веретена деления зависит по большей части от способности компонентов веретена к самоорганизации. Самоорганизация — единственный механизм сборки веретена деления в клетках лишённых центросом . Сборка биполярного веретена без участия центросом называется ацентросомальной. Она характерна для высших растений, а также наблюдается при мейозе на ранних стадиях развития некоторых животных. Более того, предполагается, что самоорганизация микротрубочек является преобладающим механизмом сборки веретена, даже в животных клетках, содержащих центросомы .
Самоорганизация веретена начинается после разрушения ядерной мембраны. Цитоплазматические микротрубочки собираются (нуклеируются) вокруг хромосом. Здесь при участии локальных стабилизирующих факторов происходит удлинение накапливающихся микротрубочек. Далее начинается организация микротрубочек с участием трёх групп моторных белков :
- Моторные белки семейства (Eg5) связываются с двумя противоположно ориентированными микротрубочками и одновременно движутся в направлении плюс-конца каждой из них. В итоге происходит сортировка антипараллельных поляризованных микротрубочек и их «сшивка» в районе плюс-конца.
- — белковые моторы семейства и , локализованные на плечах хромосом, — связывают микротрубочки находящиеся вблизи хромосом и перемещаются в направлении плюс-конца микротрубочки. Тем самым плечо хромосомы оказывается связано с плюс-концом микротрубочки, а минус-конец оказывается дистанцирован от хромосомы.
- Третья группа моторных белков перемещается в направлении минус-концов микротрубочек и обеспечивает связку минус-концов на полюсах веретена. К данной группе моторов относятся цитоплазматические динеины , . Динеин участвует в фокусировке полюсов деления совместно с многочисленными ядерными белками, например (англ. Nu clear M icrotubule- A ssociated protein 1).
Сборка с участием центросом
Во многих животных клетках, включая человеческие, в сборке веретена участвуют центросомы, являющиеся полюсами веретена деления. Также как и при сборке ацентросомального веретена, моторные и другие белки участвуют в самоорганизации микротрубочек в биполярную структуру, которая фокусируется с помощью минус-концов микротрубочек в области центросом. Центросомы при этом тоже участвуют в сборке веретена и способствуют формированию полюсов деления, но не являются неотъемлемым компонентом веретена, так как процесс сборки может протекать даже при инактивации центросом .
В зависимости от времени расхождения центросом относительно момента разрушения ядерной оболочки выделяют два механизма образования веретена :
- Если ядерная оболочка разрушается до начала расхождения центросом, то высвободившиеся хромосомы распределяются по цитоплазме, и образуется «однополюсное» веретено с микротрубочками, расходящимися от спаренных центросом. Дальнейшее образование двухполюсного веретена происходит за счёт сил отталкивания перекрывающихся микротрубочек и под действием тянущих сил астральных микротрубочек. Отталкивающее усилие между перекрывающимися микротрубочками создаётся кинезиноподобными белками Eg5. Тянущие силы, приложенные к астральным микротрубочкам, создаются цитоплазматическими динеинами, закреплёнными на внутренней поверхности клеточной мембраны.
- Второй вариант сопряжён с расхождением центросом и образованием первичного веретена до разрушения ядерной оболочки. Первичное веретено образуется за счёт тянущих сил астральных микротрубочек, которые создаются цитоплазматическими динеинами, закреплёнными на внутренней поверхности клеточной мембраны и на поверхности ядерной оболочки. Направление расхождения центросом задаётся актиновыми филаментами, которые взаимодействуют с миозином , расположенным в самих центросомах или вдоль микротрубочек. Первичное веретено является нестабильным. Для его устойчивости необходимо взаимодействие с кинетохорами хромосом и другими белками, находящимися внутри клеточного ядра.
Присоединение хромосом к веретену
Наиболее изучен механизм присоединения хромосом к веретену в животных клетках, содержащих центросомы. Во время профазы вокруг центросом формируется звёздчатая структура из микротрубочек, расходящихся в радиальном направлении. Область ядра после разрушения ядерной мембраны активно зондируется динамически нестабильными микротрубочками, которые захватываются кинетохорами хромосом. Часть хромосом быстро связывается с микротрубочками от противоположных полюсов. Другая часть хромосом сначала присоединяется к микротрубочкам исходящим от одного из полюсов. После чего перемещается в направлении соответствующего полюса. Затем связанные с одним полюсом хромосомы захватывают микротрубочки от противоположного полюса. В процессе метафазы к каждому кинетохору присоединяется порядка 10—40 микротрубочек, которые образуют кинетохорный пучок. Все хромосомы оказываются связанными с противоположными полюсами деления и собираются в метафазную пластинку в центре веретена .
Существует также альтернативная модель присоединения кинетохоров к веретену, подходящая как для клеток с центросомами, так и для клеток лишённых центросом. Согласно этой модели вблизи хромосом происходит нуклеация коротких микротрубочек при участии . Своим плюс-концом микротрубочки встраиваются в кинетохоры. Вслед за этим происходит регулируемый рост ( полимеризация ) микротрубочек. Удлиняющиеся минус-концы микротрубочек «сшиваются» и фокусируются в области полюсов деления при участии моторных белков. Центросомы (в случае их наличия) способствуют присоединению кинетохорных микротрубочек к полюсам деления .
Биполярная ориентация сестринских хроматид
Для равного распределения хромосом между дочерними клетками, важно, чтобы кинетохоры парных хроматид были присоединены к микротрубочкам, исходящим от противоположных полюсов. Нормальное биполярное прикрепление кинетохоров к противоположным полюсам называется амфителическим . Однако в процессе сборки веретена могут возникать иные прикрепления хромосом. Присоединение одного кинетохора к одному полюсу деления называется монотелическим . Присоединение сразу двух кинетохоров одной хромосомы к одному полюсу деления называется синтелическим . Возможно также и меротелическое прикрепление, при котором один кинетохор соединяется сразу с двумя полюсами .
Неверное присоединение отчасти предотвращается за счёт самой геометрии сестринских кинетохоров, которые находятся на противоположных сторонах центромерной области хромосом. К тому же неправильные прикрепления являются нестабильными и обратимыми, а нормальное биполярное крепление кинетохоров является стабильным. Стабильное соединение достигается за счёт сил натяжения, исходящих от противоположных полюсов деления. Основным компонентом регуляторной системы, ответственной за правильное присоединение кинетохоров к противоположным полюсам, является протеинкиназа aurora B .
Примечания
- , с. 506.
- , с. 508.
- , p. 1858.
- , p. 66.
- , p. 125.
- ↑ , p. 130.
- , p. 124.
- , p. 112.
- ↑ , p. 113.
- , с. 520.
- ↑ , p. 128.
- , с. 521.
- , с. 518.
- , p. 131.
- ↑ , p. 132.
Литература
- Evert R. F., Eichhorn S. E. Raven biology of plants. — 8 edition. — W. H. Freeman and Company, 2013. — 880 p. — ISBN 978-1-4292-1961-7 .
- Morgan D. O. The cell cycle: principles of control. — New science press, 2007. — 297 p. — ISBN 978-0-9539181-2-6 .
- Redei G. P. (ed.). Encyclopedia of genetics, genomics, proteomics, and informatics. — 3 edition. — Springer, 2008. — 1822 p. — ISBN 978-1-4020-6753-2 .
- Льюин Б. и др. Клетки. — М. : БИНОМ. Лаборатория знаний, 2011. — 951 с. — (Лучший зарубежный учебник). — ISBN 978-5-94774-794-2 .
- 2021-08-06
- 1