Interested Article - Нематоды

- 2021-10-07
- 2
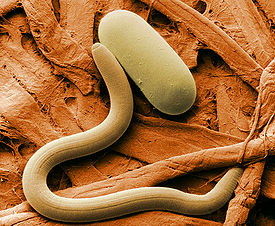
Немато́ды ( лат . Nematoda, от греч . νήμα – нить и εἶδος – форма), или кру́глые че́рви , — тип первичноротых (из группы линяющих ) беспозвоночных животных. В настоящее время описано более 24 тыс. видов паразитических и свободноживущих нематод , однако оценки реального разнообразия, основывающиеся на темпах описания новых видов (в особенности специализированных паразитов насекомых ), предполагают существование около миллиона видов .
Свободноживущие нематоды обитают в грунте водоёмов любой солёности и во всём диапазоне глубин, в обрастаниях подводных твёрдых поверхностей, в почве . Нематоды встречаются от полюсов до тропиков во всех типах сред, включая пустыни, высокие горы и глубокие моря . Они играют важную роль в экосистемах почвы и грунта водоёмов, где их численность может достигать нескольких миллионов особей на 1 м² . Нематоды Halicephalobus mephisto считаются самыми глубокоживущими сухопутными многоклеточными организмами на планете .
Многие представители стали паразитами или комменсалами животных всех крупных систематических групп, включая простейших . Они вызывают нематодозы у растений (фитонематоды) и животных (зоонематоды), в том числе человека. У поражённых растений наблюдается гибель корней, порча корнеплодов и образование галлов . Наиболее известные паразиты человека среди круглых червей: аскариды , острицы , трихинеллы , анкилостомы , ришта . Их яйца попадают в человека при несоблюдении правил личной гигиены , с загрязнённой пищей и водой. Борьба с паразитическими нематодами сводится к изгнанию их из организма хозяина. Во внешней среде производится их уничтожение медико- и ветеринарно-санитарными, а также агротехническим мероприятиями .
Ароморфозы круглых червей: первичная полость тела ( псевдоцель ), образование нервного кольца, сквозная пищеварительная система, раздельнополость и половой диморфизм .
Древнейшие достоверные находки известны из раннего мела .
Строение и физиология
Длина тела составляет от 80 мкм до 8,5 м (такую длину имеет паразит Placentonema gigantissima , живущий в плаценте кашалота ) . Свободноживущие нематоды и паразиты растений характеризуются небольшими размерами — 1–3 мм, в редких случаях свободноживущие виды достигают сантиметровой длины. Паразиты животных в большинстве своем также мелки. Самки несколько крупнее самцов . Тело нематод несегментированное, имеет нитевидную или веретеновидную, реже (у самок) бочонковидную или лимоновидную форму . В поперечном сечении тело круглое (отсюда название круглые черви ), в основе обладает билатеральной (двусторонней) симметрией тела с элементами двулучевой. Передний конец тела (голова) проявляет, кроме того, признаки трёхлучевой симметрии. За исключением некоторых органов чувств нематоды лишены жгутиковых клеток. Кровеносная и дыхательная системы отсутствуют .
Кожно-мускульный мешок
Круглые черви обладают развитым кожно-мускульным мешком . Тело покрыто гладкой или кольчатой кутикулой , под которой расположена гиподерма , или эпидермис, а под ней — тяжи продольных мышц. Кутикула, кроме того, выстилает глотку и заднюю кишку. На кутикулу приходится примерно 5–10% диаметра тела и 10–20% его объема . Самый наружный слой представляет собой тонкую эпикутикулу. Кроме эпикутикулы в состав кутикулы нематод входят еще три слоя, состоящих в основном из коллагена . Эти слои занимают основную долю кутикулы. Кортекс — самый наружный слой — залегает непосредственно под эпикутикулой. Кортекс часто разбит на кольца и может содержать особый белок — эластин . Срединный слой сильно варьирует у разных видов, он может быть гомогенным или иметь сложное строение. Внутренний, базальный, слой может быть исчерченным или слоистым. Иногда он содержит волокна, огибающие тело нематоды по спирали во взаимно противоположных направлениях. Кутикула, одевающая тело нематод, не содержит хитина , однако хитин найден в кутикуле, выстилающей глотку. По окружности тела гиподерма образует 4 валика («хорды»), вдающихся в полость тела — дорсальный (спинной), вентральный (брюшной) и два боковых. Внутри спинной и брюшной полости проходят нервные стволы, а в боковых — сенсорные нервы и выделительные каналы .
У паразитических форм эпителий может приобретать синцитиальное строение , то есть клеточных границ в нём нет, и он состоит из цитоплазматической массы с включенными в него ядрами. Ядросодержащая часть клеток эпителия соединена с поверхностным синцитиальным слоем цитоплазматическими мостиками. У паразитов в гиподерме может запасаться гликоген, необходимый для гликолиза . Под гиподермой находится слой продольных мышц, делящийся валиками гиподермы на 4 тяжа. Нематоды имеют также мышцы, связанные с репродуктивными органами и (иногда) отдельные мышечные клетки в стенке кишечника .
У представителей морского интерстициального семейства поверхность тела покрыта слоем симбиотических волокнистых синезелёных бактерий и в результате она кажется волосатой.
Полость тела

1 — передний конец тела, несущий рот; 2 — кишка ; 3 — клоака ; 4 — выделительная система ; 5 — семенник ; 6 — нервное кольцо; 7 — ; 8 — ; 9 — выделительное отверстие.
Между кожно-мускульным мешком и внутренними органами тела у более или менее крупных форм расположена первичная полость тела — псевдоцель , отличающаяся от вторичной (целома) отсутствием эпителиальной выстилки . У мелких, свободноживущих нематод полость тела мала или отсутствует совсем. У крупных паразитов животных, таких как Ascaris , она может быть очень обширна. Жидкость в полости гемоцеля находится под давлением и выполняет функции гидростатического скелета. Давление жидкости внутри тела у крупных нематод велико и составляет 75–150 мм рт. ст. (теоретически рассчитанное внутреннее давление для мелких нематод примерно в 100 раз ниже) . Через полостную жидкость также осуществляется диффузный транспорт веществ между тканям . Она содержит продукты обмена веществ, включая гемоглобин у некоторых видов, но в нем нет взвешенных клеток. У нематод имеется несколько фагоцитарных клеток, прикрепляющихся изнутри к стенкам полости тела. Эти клетки выполняют защитную функцию.
Пищеварительная система и питание
Пищеварительная система круглых червей похожа на трубку, сквозная. Она начинается ротовой полостью (стомой), переходит в глотку (обычно называемую пищеводом) , затем в переднюю, среднюю и заканчивается задней кишкой, открывающейся на заднем конце тела с брюшной стороны . Пищеварительная трубка относительно короткая и не превышает длины тела, в то время как у более высокоорганизованных животных пищеварительный тракт имеет значительно большую относительную протяженность. У паразитических нематод это во многом объясняется тем, что они питаются уже частично расщепленными продуктами, а в кишечнике червя происходит, главным образом, их усвоение. У нескольких паразитических отрядов кишка преобразована в не имеющую просвета . Ротовое отверстие терминальное или редко сдвинуто на вентральную или дорсальную сторону. Рот окружён губами, которых чаще всего бывает три , и ведёт в мускулистую глотку, образующую на своём протяжении расширения — бульбусы . Глотка имеет трёхгранный просвет, расширяющийся при сокращении радиальных мышечных волокон, и используется для засасывания пищи. Она имеет сложное строение и во многих группах хищных и паразитических нематод несёт разнообразное вооружение: у нематод, паразитирующих у животных, в глотке могут находиться кутикулярные зубы, а у паразитов растений — колющий стилет, которым паразит протыкает растительные ткани. В стенках пищевода (глотки) лежат многочисленные железы, оказывающие первоначальное воздействие на пищевой субстрат . Глотка открывается в энтодермальную по происхождению среднюю кишку, стенка которой образована лишь одним слоем высоких эпителиальных клеток, лежащих на тонкой базальной мембране. Именно в этом отделе происходит окончательное расщепление пищи и всасывание необходимых веществ, которые затем диффундируют в полостную жидкость и распространяются в ткани. В качестве запасных веществ, как правило, используется гликоген , который откладывается в валиках гиподермы. Паразитические нематоды, обитающие в бескислородной среде в полости кишечника, получают необходимое количество АТФ в результате гликолиза , для чего гликоген быстро гидролизируется . Образовавшиеся в результате неполного расщепления органические кислоты накапливаются в полостной жидкости, из-за чего она становится едкой. Пищеварительная система заканчивается задней кишкой, открывающейся у самок анальным отверстием, а у самцов — отверстием клоаки . У некоторых нематод (например, у нитчаток) анальное отверстие отсутствует, поскольку задняя кишка заканчивается слепо .
Нематоды питаются в основном бактериями, водорослями, детритом ; есть среди них и хищники, нападающие на мелких многоклеточных, в том числе и на других нематод (нематода мононх - Mononchus , названная так потому, что имеет в ротовой полости один большой зуб, живет около 18 недель и за это время съедает до 1 тыс. нематод других видов), многие — паразиты животных, грибов и растений .
Выделительная система
Состоит из одноклеточных кожных (гиподермальных) желез, заменивших протонефридии , которые исчезли в связи с утратой нематодами ресничных образований. Обычно имеется одна шейная железа. У части свободноживущих нематод она массивна и имеет короткий выделительный проток (её дополняют более мелкие железы, лежащие по бокам тела). У большинства почвенных и паразитических нематод шейная железа связана с длинными экскреторными каналами, расположенными в боковых утолщениях гиподермы. У лошадиной аскариды, достигающей 40 см длины, такая выделительная система образована одной гигантской клеткой.
Через боковые каналы выводятся вырабатываемые в теле жидкие продукты выделения. У нематод имеются также особые фагоцитарные органы (фагоцитарные клетки), в которых задерживаются и накапливаются различные нерастворимые продукты обмена веществ и посторонние тела, проникающие в организм, например бактерии. Следовательно, нерастворимые частицы, накопившиеся в фагоцитарных органах, не удаляются из тела, а только устраняются таким образом из жизненного круговорота организма. Такие органы называются «почками накопления»; они имеют вид крупных звездчатых клеток. Они расположены в полости тела по ходу боковых выделительных каналов, в передней трети тела. Фагоцитарных клеток у нематод четыре, но могут встречаться особи с меньшим (2–3) или большим (до 6) числом .
Нервная система и органы чувств
Нервная система состоит из окологлоточного нервного кольца и нескольких продольных нервов. Нервное кольцо находится на уровне середины глотки и наклонено дорсальным краем вперёд (в некоторых группах наклон обратный). По своему строению нервное кольцо является единым круговым ганглием и, видимо, выполняет роль основного ассоциативного органа. От него берут начало вентральный нервный ствол и дорсальный нерв, остальные продольные нервы с ним непосредственно не связаны. Вентральный нервный ствол содержит тела нейронов, другие продольные нервы не имеют тел и являются пучками отростков нейронов вентрального ствола. Все продольные стволы проходят интраэпителиально — в валиках гиподермы.
Органы чувств представлены многочисленными сенсиллами : осязательными , губными (сосочками), органами самцов, обонятельными и сенсорно-железистыми органами — . На заднем конце тела свободноживущих нематод имеются терминальные хвостовые железы, секрет которых служит для прикрепления к субстрату . Эти органы чувств являются механо- , хемо- или реже фоторецепторами или обладают смешанной чувствительностью и всегда ассоциированы с железистыми клетками. Основными органами дальней химической рецепции являются амфиды — сложно устроенные парные органы на переднем конце тела, имеющие разнообразную форму. К другим органам чувств головы относятся головные сенсиллы, подчинённые в своём расположении радиальной симметрии и расположенные в три или два ряда. У некоторых свободноживущих нематод, кроме того, обнаружены внутренние механорецепторы — .
Американские учёные, проводя эксперименты над нематодами, обнаружили, что те способны к обучению на собственном опыте. Взрослые черви, почуяв запах пищи, сразу же ползли на него кратчайшим путём, в то время как молодые долго сомневались и даже не всегда отправлялись на поиски. Кроме того, у опытных нематод в ответ на запах активировались три парных нейрона — чего не происходило с молодыми .
Дыхание
Специализированных органов дыхания у нематод нет, многие виды (обитатели гниющих сред, паразиты кишечника животных) являются анаэробами . Такие паразиты кишечника, как Ancylostoma и Necator питаются кровью, при этом они используют кровь не только как пищевой субстрат, но и для обеспечения себя кислородом: стенки их кишечника поглощают кислород из проглоченной крови, отщепляя его от гемоглобина.
Половая система
В подавляющем большинстве нематоды имеют отчётливый внешний половой диморфизм и раздельнополы, в очень редких случаях у нематод наблюдается гермафродитизм или способность к партеногенезу . Нематоды откладывают яйца, реже живородящи. Из оплодотворённых яиц вылупляются личинки . Это происходит во внешней среде или ещё в половых путях самки (яйцеживорождение) . У самцов задний конец тела загнут на брюшную сторону (у некоторых видов задний конец самца образует две расширенные лопасти ), и имеется сложный копулятивный аппарат. Роль удержания самки во время копуляции играют разнообразные и (у рабдитидных нематод) бурсы. Спермии вводятся с помощью спикул, выдвигающихся из клоакального отверстия и возвращающихся обратно благодаря мышцам . Внутренние половые органы в исходном варианте парные и имеют трубчатое строение. У самок имеется единичный или двойной набор из яичника, яйцевода и матки; влагалище всегда единственное. Участок половой трубки самки может специализироваться как семяприемник, где хранятся сперматозоиды и происходит оплодотворение . У самцов имеются один или два семенника с семяпроводами и непарный семяизвергательный канал. Спермии нематод имеют крайне разнообразное строение, лишены жгутиков и обладают амебоидной (но не актиновой ) подвижностью .
Оплодотворение внутреннее. Для самок некоторых видов отмечено выделение феромонов — веществ, привлекающих самцов. В ходе совокупления задний конец тела самца закручивается вокруг тела самки, так что отверстие клоаки самца плотно прижимается к половому отверстию самки. Самец вводит в женское половое отверстие спикулы и расширяет его, обеспечивая передачу спермы во влагалище. Оплодотворение происходит в семяприемнике. Оплодотворенное яйцо покрывается многослойной оболочкой и выводится из организма самки. Развитие яйца начинается в половых путях самки, так что отложенное яйцо содержит зародыш на разных стадиях развития. Свободноживущие нематоды откладывают несколько десятков яиц. Количество яиц у зоопаразитических видов всегда велико, но зависит от размера животного (миллиарды яиц откладывает самая крупная нематода Placentonema gigantissima ). Для некоторых нематод (например, для трихинелл , ришт ) характерно живорождение.
У большинства нематод, как и у многих других животных, пол определяется генетически, при помощи половых хромосом . Если в оплодотворённом яйце две Х-хромосомы, из яйца разовьётся самка, если одна — самец. Однако у двух наиболее изученных видов нематод, Caenorbabditis elegans и , особи с двумя Х-хромосомами не самки, а гермафродиты. Их гонады на поздних личиночных стадиях производят спермии. У гермафродитов спермии поздних личинок переползают в специальные хранилища — сперматеки — и здесь ждут своего часа. Половые железы взрослых гермафродитов производят уже не спермии, а яйцеклетки. Они могут быть оплодотворены как собственными спермиями из сперматеки, так и спермиями самца в результате спаривания. Предки C. elegans и C. briggsae были нормальными раздельнополыми червями. Это следует, в частности, из того, что все прочие виды рода Caenorhabditis — раздельнополые. По-видимому, гермафродитизм у C. elegans и C. briggsae является «эволюционно молодым», новым признаком.
Развитие
Жизненный цикл включает стадию яйца, четыре ювенильные стадии и стадию взрослой нематоды. Развитие прямое, и молодые особи имеют почти все органы уже в момент выхода из яйца, за исключением репродуктивной системы. Рост на протяжении четырех ювенильных и одной взрослой стадии сопровождается четырьмя линьками. У некоторых видов первые две из них могут происходить под оболочкой яйца до вылупления. Животное становится взрослым после четырех линек. Больше нематоды не линяют, но их рост может продолжаться и после четвертой линьки. Рост у нематод сопровождается не только изменением размеров и массы тела, но и изменением пропорций и существенными перестройками внутренней организации. Различные отделы тела растут с разной скоростью.
Наличие прямого развития у нематод — следствие исходно мелких размеров представителей этой группы. Развитие с пелагической личинкой, предоставляющее организмам значительные преимущества в смысле широкого расселения и позволяющее им использовать пищевые ресурсы пелагических биотопов, сопряжено с массовой гибелью личинок. Поэтому пелагическое развитие доступно только относительно крупным формам, продуцирующим большое количество яиц. Мелкие формы, обладающие органической плодовитостью, как правило, вынуждены переходить к прямому развитию.
Для части зоопаразитических нематод в ходе раннего развития зарегистрировано явление диминуции хроматина . Оно заключается в том, что соматические клетки на третьем шаге дробления теряют около 85% своей ДНК . Только клетки зародышевой линии сохраняют полный геном .
Эутелия
Нематоды демонстрируют эутелию. Клеточные деления (исключая клетки гонад) прекращаются незадолго до конца эмбрионального развития. Таким образом, число клеток взрослого животного постоянно и характерно для вида. (Эутелия характерна далеко не для всех нематод. Большинство свободноживущих морских нематод не обладают постоянством клеточного состава.) Различные органы и ткани также содержат фиксированное число клеток, большинство из которых имеется уже ко времени вылупления молодых особей из яйца. Так, например, взрослая особь Rhabditis имеет 200 нервных клеток, 120 эпидермальных клеток и 172 клетки пищеварительного канала. Гермафродитные особи Caenorhabditis elegans всегда имеют 959 соматических ядер. Только клетки зародышевой линии продолжают делиться после наступления взрослого состояния и их число различно. Поскольку увеличения числа клеток во время постэмбрионального развития не происходит, рост нематод по большей части осуществляется за счет увеличения размеров клеток.
Локомоция
Большинство нематод двигаются вперёд и в обратном направлении, используя синусоидальные , волнообразные движения тела. Эти движения происходят в дорсовентральной плоскости, что достигается за счёт попеременных сокращений спинных и брюшных продольных мышц. Изъятые из естественного субстрата нематоды изгибаются, но не могут направленно двигаться вперёд. Эффективное волнообразное движение требует наличия субстрата, такого как гранулы песка, или поверхностной пленки жидкости, что позволяет нематодам отталкиваться. В пелагических условиях такое движение малоэффективно. Большинство свободноживущих нематод являются интерстициальными животными, они быстро и эффективно движутся, отталкиваясь от стенок узких пространств, в которых живут. Размер пространства, позволяющего совершать оптимальные волнообразные движения, составляет около 1,5 диаметра червя, и для большинства почвенных нематод размер почвенных пор от 15 до 45 мкм идеален.
Хвостовая железа (спинеретта) типична для многих свободноживущих нематод, включая большинство морских видов. Она открывается на заднем конце тела, который оттянут в виде конического хвоста. Для некоторых нематод характерна система двух типов желез (один для прикрепления, другой для открепления). Железа используется для временного прикрепления к субстрату. Такое прикрепление позволяет быстро подтянуть тело к субстрату, избегая опасности.
Филогения
Палеонтологический метод как способ проведения филогенетического исследования мало применим к нематодам, так как эти организмы, во-первых, очень мелкие, что делает их обнаружение крайне трудным и, во-вторых, что более важно, у них мало твёрдых структур, такие как кости или панцирь, которые подходят для различных процессов окаменения. Таким образом, традиционно систематика нематод основывалась почти исключительно на таксономической классификации, т. е. на морфологических сходствах современных особей. В эволюционных представлениях было и остается много сомнений, вызванных тем, что ископаемые остатки нематод практически отсутствуют. Однако имеется несколько окаменелых нематод, которые более или менее хорошо сохранились в янтаре , но это относительно «молодые» формы, существовавшие только от нескольких тысяч до нескольких миллионов лет назад. Большая часть этих особей представляет собой специализированных паразитов насекомых, а не примитивные древние формы. Даже самая старая ископаемая нематода является паразитом насекомых из отряда Mermithida . Эта особь была обнаружена в ливанском янтаре мелового периода и имеет возраст не более 130 млн лет. Следовательно, эта нематода тоже слишком молода для установления основных эволюционных событий, потому что у неё есть уже все необходимые признаки отряда, семейства и рода. Описанная как Heleidomermis libani , она просто представляет собой новый вид в дополнение к известным современным видам рода. Тем не менее, это уже очень важное открытие, показывающее, что нематоды являются крайне древней группой, которая эволюционирует, по-видимому, очень медленно.
Изучение современного географического распространения нематод в связи с ходом таких естественных геологических событий, как тектоника плит и материковый дрейф, может дать достоверные представления об эволюции нематод за последние 300 млн лет. Это особенно верно, когда принимаются в расчет результаты современной сравнительной морфологии. Там, где есть достаточно информации, филогенетическая система, основанная на морфологических признаках, хорошо согласуется с данными биогеографии. Кроме того, эти сведения подтверждаются генетическими данными молекулярно-таксономических исследований. 300 млн лет назад многие из существующих ныне отрядов нематод уже были. К этому времени современная форма тела нематод, таким образом, была в значительной степени достигнута, как следует из относительно незначительных морфологических изменений, которые произошли после этого. Следовательно, как группа нематоды намного старше, чем 300 млн лет, а их основная морфологическая эволюция произошла ещё раньше.
Когда провели сравнение состава цитохрома с и некоторых гистонов у свободноживущей нематоды-бактериофага Caenorhabditis elegans , наиболее широко используемого модельного организма для изучения физиологии , эволюции и генетики , с таковым у млекопитающих , подсчитали, что время их разделения составляет более 1 млрд лет. Эти данные указывают на то, что нематоды существовали уже 1 млрд лет назад, и хорошо согласуются с вышеупомянутыми свидетельствами того, что нематоды на самом деле древняя группа. Среди нематод наиболее древним таксоном является отряд Enoplida .
Исследования над морскими гастротрихами (отряд ) позволяют подтвердить и развить ранее высказывавшийся взгляд о близости нематод к данной группе животных. Черты сходства касаются очень многих признаков. У представителей отряда Macrodasyoidea имеется хвост, лежащий позади расположенного на брюхе ануса, терминальное ротовое отверстие, наблюдается сходное строение ротовой полости, пищевода и кишки. Пищевод гастротрих имеет радиальную мускулатуру и трехгранный просвет, как у нематод. Кишка и у тех, и у других состоит из одного слоя энтодермальных клеток. Очень сходны ресничные ямки некоторых морских гастротрих с амфидами нематод. Велико сходство некоторых физиологических особенностей, закономерностей роста и и др. Вместе с тем гастротрихи более примитивны, чем нематоды, менее специализированы. Во многих чертах организации (ресничный покров, обонятельные ресничные ямки, строение полового аппарата и другие признаки) гастротрихи близки к турбелляриям . Наиболее близко к истине представление, согласно которому гастротрихи отчленились от турбеллярий (прямокишечных) и в свою очередь от основания их ствола отошли нематоды.
Систематика

Впервые группа была определена Карлом Асмундом Рудольфи в 1808 году под именем Nematoidea . Позже была последовательно классифицирована как семейство Nematodes Бурмистром в 1837 году и как порядок Nematoda в 1861 году .
Таксономический статус и систематическое положение некоторых групп нематод (классов, подклассов, отрядов) дискутируется . Например, по некоторым старым классификациям крупнейший класс Chromadorea принимали в узком объёме (без Ascaridida , Spirurida , Tylenchida ) и включали в ранге отряда Chromadorida Chitwood, 1933 в состав сборного парафилетического подкласса нематод Аденофореи, или афазмидиевые (Adenophorea, Aphasmidia Chitwood et Chitwood, 1933 ) . Впервые деление нематод на два подкласса в 1930-х годах обосновал Б. Читвуд (Chitwood B., 1933, 1937). Однако, предложенное им название Phasmidia Chitwood et Chitwood, 1933 оказалось уже занято более старым сходным названием одного из отрядов насекомых. В большинстве работ XX века нематодологи использовали такие названия подклассов :
- Подкласс Adenophorea Linstow, 1905 (или Aphasmidia Chitwood et Chitwood, 1933 ) с отрядами Chromadorida (позднее разделённого на отряды Araeolaimida , Desmodorida , Desmoscolecida , Monhysterida ) и Enoplida ( Dorylaimida , Mermithida , Muspiceida , Trichocephalida )
- Подкласс Secernentea Linstow, 1905 (или Phasmidia Chitwood et Chitwood, 1933 ) первоначально только с двумя отрядами Rhabditida и Spirurida (затем они были раздроблены на , Ascaridida , , Diplogasterida , , Rhabditida , Spirurida , Strongylida , Tylenchida )
Искусственность такого классического взгляда на систематику нематод (особенно Adenophorea) всё более подтверждали новые анатомические и молекулярно-генетические исследования. В некоторых более поздних системах, где все нематоды рассматриваются в ранге класса, выделяют 3 подкласса (Малахов, 1986) .
- Подкласс Enoplea ( Dorylaimida , Enoplida , Marimermithida , Mermithida , Mononchida , Trichocephalida )
- Подкласс Chromadoria ( Araeolaimida , Chromadorida , Desmoscolecida , Desmodorida , Monhysterida , Plectida )
- Подкласс Rhabditia ( Ascaridida , , Rhabditida , Spirurida , Strongylida , Tylenchida )
Современная классификация
По данным на 2011 год, тип нематод включает 3 класса, 31 отряд, 267 семейств, 2829 родов и 24 783 вида, причём ископаемые таксоны представлены в 2 родах 10 видами; есть также 7 родов и 7 видов, известных только в ископаемом состоянии (ранее высказывались оценки от 15 до 80 тысяч , а реальное разнообразие нематод — с учётом перспектив описания новых видов — оценивается в миллион видов ).
Тип включает три класса и около 30 отрядов :
-
Тип
Nematoda
-
Класс
Chromadorea
- Отряд Benthimermithida
- Отряд Chromadorida
- Отряд Desmodorida
- Отряд Desmoscolecida
- Отряд Diplogasterida
- Отряд
- Отряд
- Отряд Monhysterida (вкл. Araeolaimida )
- Отряд (вкл. Tylenchina )
- Отряд Plectida
- Отряд Rhabditida , включает большинство крупных паразитов позвоночных , а также модельную нематоду Caenorhabditis elegans
- Отряд
- Отряд Spirurida (вкл. Ascaridina )
- Отряд
-
Класс
Enoplea
- Отряд Alaimida (или подотряд Alaimina в отряде Enoplida )
- Отряд Enoplida
- Отряд Ironida (или подотряд Ironina в отряде Enoplida )
- Отряд Isolaimiida
- Отряд Oncholaimida (или подотряд Oncholaimina в отряде Enoplida )
- Отряд Rhaptothyreida
- Отряд
- Отряд Trefusiida (или подотряд Trefusiina в отряде Enoplida )
- Отряд Triplonchida
- Отряд Tripylida
- Отряд Tripyloidida (или подотряд Tripyloidina в отряде Enoplida )
-
Класс
(или подкласс в
Enoplea
)
- Отряд (или подотряд и в отряде Mononchida )
- Отряд Dioctophymatida — как правило, крупные нематоды, паразитирующие в кишечнике, желудке и почках млекопитающих и птиц
- Отряд Dorylaimida — свободно движущиеся морские, пресноводные и почвенные нематоды
- Отряд Marimermithida
- Отряд Mermithida (в том числе паразиты беспозвоночных из семейства Mermithidae )
- Отряд Mononchida
- Отряд Muspiceida
- Отряд Trichocephalida — паразиты позвоночных животных (включая широко распространённых паразитов из родов трихинелла и власоглав )
-
Класс
Chromadorea
Нематодозы
Нематодозы — паразитарные болезни ( гельминтозы ) человека, животных и растений, вызываемые нематодами. Среди нематодозов выделяют две группы: (когда развитие яиц и/или личиночных форм происходит в почве , воде или на предметах домашнего обихода) и бионематозы (когда цикл развития связан со сменой хозяев и передача возбудителей осуществляется переносчиками — комарами , мошками , слепнями ). Сохранение и развитие яиц и личинок нематод во внешней среде возможно лишь при определённых температурных условиях, достаточной влажности и доступе кислорода .
Нематодозы у человека
Наряду с другими гельминтозами, нематодозы имеют наибольшее значение в патологии человека. Паразиты локализуются в большинстве тканей и органов человека ( желудочно-кишечный тракт , мышцы , органы дыхания , печень , почки и т. д.). Обычно заражение человека происходит при заглатывании зрелых (инвазионных) яиц или личинок нематод с частицами почвы, водой, продуктами питания. На территории стран бывшего СССР у человека встречаются (зарегистрированы): аскаридоз , энтеробиоз , трихоцефалез , трихинеллёз и другие нематодозы . Терапия (лечение) большинства нематодозов у человека при своевременном обращении за медицинской помощью не представляет трудностей .
Нематодозы у животных
Нематодозы встречаются (описаны) у всех видов позвоночных животных . Большая часть видов нематод паразитирует в пищеварительном тракте . На распространённость нематодозов среди животных влияют: климатические условия; наличие и количество промежуточных хозяев; условия, в которых содержатся животные; качество и своевременность проводимых лечебных и профилактических мероприятий. Наибольший экономический ущерб животноводству наносят аскаридоз , , , , , трихинеллёз и ряд других нематодозов .
Нематодозы у растений
Нематодные болезни растений (травянистых, древесных, кустарников) вызывает ряд вредоносных растениеядных нематод. Встречаются у многих диких и культурных растений. Наиболее часто внешние признаки нематодных поражений растений проявляются замедлением появления всходов, роста и развития саженцев, слабым цветением, частичной (иногда значительной) гибелью растений в молодом возрасте, снижением или гибелью урожая. В процессе питания нематоды нарушают целостность корней, тем самым способствуя проникновению в растение патогенных грибов, бактерий и вирусов . Внедрение нематод в корни растений обычно вызывает сильное ветвление корневой системы и отгнивание мелких корней (свекловичная, картофельная , овсяная гетеродеры), образование галлов разной формы (галловые нематоды на корнях овощных и технических культур), заострённых вздутий — «клювиков» (угрица — лат. Anguina radicicola — на корнях диких злаков), язв, приводящих к отмиранию корней. Стеблевые нематоды вызывают веретеновидное утолщение стеблей, недоразвитие листовой пластинки и её деформацию, у земляники : вздутие листовых черешков, усов и гофрировку листовой пластинки; образование на периферии клубней картофеля мягких тёмных пятен; растрескивание донца и разрыхление ткани сочных чешуй у луковичных растений .
Всё разнообразие направлений, методов и средств защиты сельскохозяйственных культур и других полезных человеку растений от нематод сводится к следующему:
- организационные профилактические мероприятия, предотвращающие распространение фитопатогенных нематод с семенами, посадочным и тому подобным материалом;
- физико-химические средства и методы уничтожения нематод в почве, в зараженных растениях и в их отдельных органах;
- агротехнические мероприятия;
- биологические средства и методы защиты культурных растений от нематод.
Значение нематод в природе
Огромное число видов нематод обитают на дне водоемов и участвуют в переработке органических остатков, т. е. представляют важный комплекс сапрофагов в морских и пресноводных биоценозах. Почвенные нематоды играют существенную роль в почвообразовании. Среди свободноживущих нематод имеются виды, питающиеся бактериальной флорой, грибами. Широкий трофический спектр нематод свидетельствует о существенном значении их как консументов и редуцентов в биоценозах.
Нематоды — паразиты растений и животных в природных ценозах представляют мощный фактор регуляции численности поражаемых ими видов и фактор естественного отбора в природе.
Живучесть
Российские биологи взяли более 300 образцов замерзшей почвы в арктической части Сибири и исследовали их, после чего в почве им удалось обнаружить двух нематод разных видов. Исследователи поместили их в чашку Петри с питательной средой при температуре около 20°C, после чего нематоды начали проявлять признаки жизни. Нематоды одного из видов, , были в образцах почвы возрастом 32 тыс. лет. Нематоды второго вида, , были в образце, радиоуглеродное датирование которого показало, что его возраст еще больше — 42 тыс. лет. Таким образом, нематоды поставили рекорд пребывания в замороженном состоянии ( анабиозе ) среди животных .
Сотни нематод вида Caenorhabditis elegans пережили катастрофу шаттла «Колумбия» в 2003 году.
В культуре
Нематоды упоминаются в фильме Хеллбой: Герой из пекла (2004).
Примечания
- ↑ Hodda, Mike. (англ.) // Zootaxa . — 2011. — No. 3148 . — P. 63—95 . 28 апреля 2021 года.
- ↑ Lambshead P. J. D. Recent developments in marine benthic biodiversity research // Oceanis. — 1993. — Vol. 19 (6). — P. 5—24.
- ↑ : [ 26 августа 2021 ] / А. В. Чесунов // Нанонаука — Николай Кавасила. — М. : Большая российская энциклопедия, 2013. — С. 353. — ( Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов ; 2004—2017, т. 22). — ISBN 978-5-85270-358-3 .
- ↑ Эдвард Э. Рупперт, Ричард С. Фокс, Роберт Д. Барнс. Зоология беспозвоночных. Том 4. — 2008.
- . Дата обращения: 13 июня 2011. 20 сентября 2011 года.
- ↑ .
- ↑ Нематоды / Г. А. Платонова // Моршин — Никиш. — М. : Советская энциклопедия, 1974. — ( Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров ; 1969—1978, т. 17).
- Poinar G.O. et al. // Fundam. appl. Nemawl. — 1994. — № 475—477 .
- Губанов Н. М. // Доклады АН СССР. — 1951. — Т. 77 , № 6 . — С. 1123—1125 . 16 января 2024 года.
- ↑ Языкова И.М. Зоология беспозвоночных. Курс лекций. — 2011.
- Нематоды // Биология. Современная иллюстрированная энциклопедия / Гл. ред. А. П. Горкин. — М. : Росмэн, 2006. — 560 с. — (Современная иллюстрированная энциклопедия). — ISBN 5-353-02413-3 .
- ↑ .
- ↑ Г. Л. Билич, В. А. Крыжановский. Биология. Полный курс. Зоология. — 2002.
- ↑ . Дата обращения: 18 сентября 2021. 18 сентября 2021 года.
- ↑ В. А. Догель. Зоология беспозвоночных. — 1981.
- . Портал "Чердак: наука, технологии, будущее" . chrdk.ru. Дата обращения: 10 января 2017. 11 января 2017 года.
- Smith H. // WormBook. — 2006. — doi : . 2 апреля 2016 года.
- Юшин В.В., Малахов В.В. // Биология моря. — 2014. — № 40 . — P. 83–94. 2 июня 2018 года.
- А. Марков, Е. Наймарк. Эволюция. Классические идеи в свете новых открытий. — 2014.
- ↑ В. В. Малахов. Нематоды. Строение, развитие, система и филогения. — 1986.
- ↑ Б. Вайшер, Д. Д. Ф. Браун. Знакомство с нематодами. Общая нематология. — 2001.
- . www.zoofirma.ru . Дата обращения: 18 августа 2022. 18 августа 2022 года.
- ↑ , 1957, Дата обращения: 22 февраля 2011. 7 июля 2012 года.
- ↑ .
- Определитель паразитических нематод. Т. 1. Спирураты и филяриаты / Под ред. К. И. Скрябина . — М. — Л. : Изд-во АН СССР, 1949. — 520 с. — С. 243.
- Зоология беспозвоночных. Т. 1: От простейших до моллюсков и артропод / Под ред. В. Вестхайде и Р. Ригера. — М. : КМК, 2008. — iv + 512 с. — ISBN 978-5-87317-491-1 .
- Margulis L., Chapman M. J. . Kingdoms & Domains: An Illustrated Guide to the Phyla of Life on Earth. 4th ed. — Amsterdam: Academic Press, 2009. — 659 p. — ISBN 978-0-12-373621-5 . — P. 274.
- (англ.) . ucdavis.edu. Дата обращения: 7 августа 2017. 29 июля 2017 года.
- ↑ Нематодозы — статья из Большой советской энциклопедии .
- Нематодозы // 1. Малая медицинская энциклопедия. — Медицинская энциклопедия. 1991—96 гг. 2. Первая медицинская помощь. — Большая Российская Энциклопедия. 1994 г. 3. Энциклопедический словарь медицинских терминов. — Советская энциклопедия. — 1982—1984 гг . — М.
- Нематодные болезни растений — статья из Большой советской энциклопедии . Свешникова Н. М..
- К. Л. Даддингтон. Хищные грибы — друзья человека. — 1959.
- ↑ Шарова И. Х. Зоология беспозвоночных.
- . Дата обращения: 1 августа 2023. 31 июля 2023 года.
- . 2003-05-02. из оригинала 18 сентября 2021 . Дата обращения: 18 сентября 2021 .
Литература
- Вайшер Б., Браун Д. Д. Ф. . — София-М., 2001.
- Нематоды // Моршин — Никиш. — М. : Советская энциклопедия, 1974. — ( Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров ; 1969—1978, т. 17).
- Нематоды // Биологический энциклопедический словарь. 2-е изд / Гл. ред. М. С. Гиляров; Редкол.: А. А. Бабаев, Г. Г. Винберг, Г. А. Заварзин и др. — М. : Сов. энциклопедия, 1986.
- Нематоды // Энциклопедия Кольера. — Открытое общество, 2000.
- Ерошенко А. С., Волкова Т. В. . Нематоды растений Дальнего Востока России: Отряды Tylenchida и Aphelenchida. — Владивосток: Дальнаука, 2005. — 225 с. — ISBN 5-8944-0602-7 .
- Малахов В. В. / Отв. ред. О. И. Белогуров ; АН СССР, Дальневосточный науч. центр, Ин-т биологии моря. — М. : Наука , 1986. — 216 с.
- Рупперт Э. Э., Фокс Р. С., Барнс Р. Д. . Зоология беспозвоночных: функциональные и эволюционные аспекты. Т. 4: Циклонейралии, Щупальцевые и Вторичноротые. — М. : Издат. центр «Академия», 2008. — 352 с. — ISBN 978-5-7695-3497-3 .
- Чесунов А. В. . Биология морских нематод. — М. : Товарищество научных изданий КМК, 2006. — 367 с. — ISBN 5-87317-325-7 .
- Гагарин В. Г. . — Л. , 1981.
- Гагарин В. Г. . — М. , 2001.
- Зиновьева С.В. и др. . — М. , 2012.
- De Ley P. & Blaxter M.L. Systematic position and phylogeny // / Donald L. Lee (Ed.). — London: Taylor and Francis, 2010. — P. 1—30. — 635 p. — ISBN 0415272114 .
- Andreas Schmidt-Rhaesa. . — Berlin/Boston: Walter de Gruyter, 2014. — 759 p. — ISBN 978-3-11-027425-7 .
- Maggenti, Armand R. / Mortimer P. Starr (ed.). — New York, Heidelberg, Berlin: Springer-Verlag, 1981. — 372 p. — ISBN 978-1-4612-5940-4 .
- Maggenti, Armand R. Nemata: Higher Classification // / Nickle William R. (ed.). — Marcell Dekker Inc (Taylor & Francis, 2015), 1991. — P. 147—187. — 1043 p. — ISBN 978-3-11-027425-7 .
Ссылки
- . Дата обращения: 7 августа 2012. 2 июня 2013 года.
- . Дата обращения: 7 августа 2012. 4 февраля 2013 года.
- . Дата обращения: 7 августа 2012. 12 октября 2013 года.
|
Первичноротые
(Protostomia)
|
|||||||
|---|---|---|---|---|---|---|---|
| Спиральные |
|
||||||
| Линяющие |
|
||||||
|
|||||||
- 2021-10-07
- 2