Гаплогруппа R (Y-ДНК)
- 1 year ago
- 0
- 0

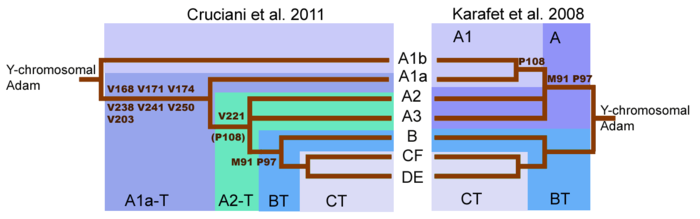
В популяционной генетике и геногеографии человека, изучающих Y-хромосомные гаплогруппы, гаплогруппа P-P295 (она же K2b2 ) — патрилинейная наследственность , возникшая, вероятнее всего, более 30 тысяч лет назад в Азии, определяемая по целому ряду уникальных меток (а именно: 92R7_1, 92R7_2, L138, L268, L405, L471/PF5989, L536/PF5860, L721/PF6020, L741, L768/PF5976/YSC0000274, L779/PF5907/YSC0000251, L781/PF5875/YSC0000255, M45 /PF5962, M74/N12, P27.1_1/P207, P27.1_2, P69, P226/PF5879, P228/PF5927, P230/PF5925, P235/PF5946, P237/PF5873, P239, P240/PF5897, P243/PF5874, P244, P281/PF5941, P282/PF5932, P283/PF5966, P284, P295/PF5866/S8, Page83, V231), что может говорить о периоде достаточно долгой её изоляции. Поскольку от неё через P1-M45=K2b2a происходят гаплогруппы R и Q , является макрогруппой .
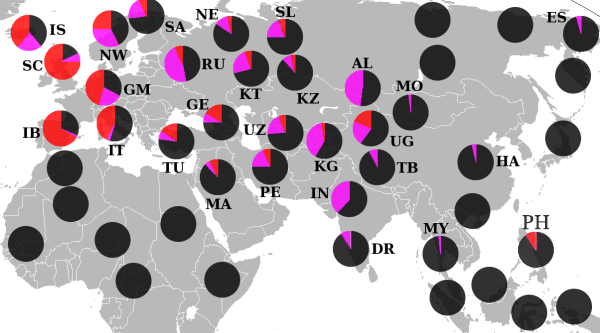
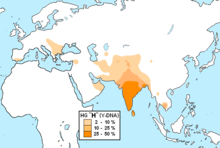
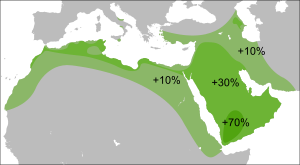
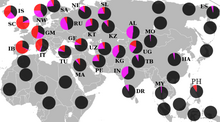
Потомки гаплогруппы P =K2b2 доминируют среди индоевропейцев (подветвь R ) и тюркских народов (подветвь R и Q ), и американских индейцев (подветвь Q ), а также составляют значительную часть населения Центральной и Южной Азии .
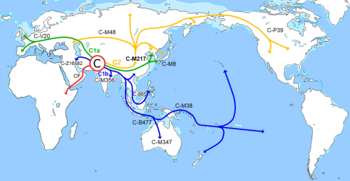
Гаплогруппа P (она же K2b2) произошла от субклады K2b-M1221/P331/PF5911 примерно 44 300 л. н. Последний общий предок современных носителей гаплогруппы P жил 31 900 лет назад (даты определены по снипам компанией YFull ). Субклада K2b произошла от гаплогруппы K2=K(xLT) (M526), которая в свою очередь происходит от гаплогруппы K (M9). От гаплогруппы P=K2b2 произошла субклада P1=K2b2a (M45).
Древним носителем предковой K2b* был , живший ~40 тыс.л.н. недалеко от Пекина . А вот пришли ли его предки северным континентальным путём (через Алтай) или южным береговым (через Индокитай и Китай), и как именно расселялись потомки – на этот счёт в ходу две гипотезы:
Возможно, что большинство или многие из описанных случаев P* на Востоке в действительности относятся к недавно открытой кладе P2, тогда группа распадается на "сибирский кластер" P1 и "филиппинский кластер" P2. Но в любом случае, присутствие в китайском регионе P* и MS оставляет хорошую поддержку версии о возникновении группы P именно где-то на юго-востоке Азии. С другой стороны, родственная группа NO1 (K2a2) , хоть и подверглась наибольшей дивергенции в монгольском регионе, самые древние линии N* и O* также в Сычуани, а её предковая K2a* – у Усть-Ишимского человека и (K2a1) на древних Карпатах и (единичные случаи) в Индии. Так что возможен почти любой сценарий перемещения групп в эпоху 40-30 тыс.л.н. между Восточной Азией и Сибирью: это было время огромной мобильности маленьких популяций.
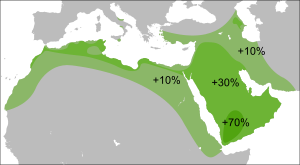
Подавляющее большинство носителей M45 относятся к одной из двух её больших ветвей: R или Q , однако парагруппа P* также встречается у сибирских и дальневосточных народов, в Средней и Восточной Азии , а также на хорватском острове Хвар (что могло произойти вследствие миграции аваров ), у ашкеназов , индейцев и у народов Южной Азии и Океании . У филиппинского племени аэта базальная гаплогруппа P* достигает 28%, обнаружены субклады P1* (М45) и P2 (Р-B253). У тиморцев Индонезии базальная P* достигает 10%. Остров Лусон является также единственным местом где ветви Р*, Р1* и очень редкая Р2 встречаются вместе , а также высока доля гаплогруппы K2b1 .
| Эволюционное древо гаплогрупп Y-хромосомы человека | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||||||||||||||||||||||||||
| Y-хромосомный Адам | |||||||||||||||||||||||||||||||||
| A0-T | |||||||||||||||||||||||||||||||||
| A00 | A0 | A1 | |||||||||||||||||||||||||||||||
| A1a | |||||||||||||||||||||||||||||||||
| A1b1 | BT | ||||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||||
| DE | CF | ||||||||||||||||||||||||||||||||
| D | E | C | F | ||||||||||||||||||||||||||||||
| F1 F2 F3 | |||||||||||||||||||||||||||||||||
| G | |||||||||||||||||||||||||||||||||
| H | IJK | ||||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||||
| I | J | K2 | |||||||||||||||||||||||||||||||
| L (K1a) | T (K1b) | K2a/K2a1/ NO /NO1 | |||||||||||||||||||||||||||||||
| N | O | /P1 | |||||||||||||||||||||||||||||||
| S (K2b1a) | M (K2b1b) | Q | R | ||||||||||||||||||||||||||||||
| Ключевые понятия |

|
|
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
|
|
В статье
не хватает
ссылок на источники
(см.
рекомендации по поиску
).
|
| Гаплогруппа P | |
|---|---|
| Тип | Y-ДНК |
| Время появления | ~32 тыс. л. н. (41–27 тыс.л.н. ) |
| Место появления | Центральная Азия , Сибирь или Китай |
| Предковая группа | K2b |
| Сестринские группы | K2b1 (далее M и S ) |
| Субклады | P1 (далее Q и R ), P2 |
| Мутации-маркеры | 92R7_1, 92R7_2, L138, L268, L405, L471/PF5989, L536/PF5860, L721/PF6020, L741, L768/PF5976/YSC0000274, L779/PF5907/YSC0000251, L781/PF5875/YSC0000255, M45 /PF5962, M74/N12, P27.1_1/P207, P27.1_2, P69, P226/PF5879, P228/PF5927, P230/PF5925, P235/PF5946, P237/PF5873, P239, P240/PF5897, P243/PF5874, P244, P281/PF5941, P282/PF5932, P283/PF5966, P284, P295/PF5866/S8, Page83, V231 |