
Педи (народ)
- 1 year ago
- 0
- 0

Управляемые циклическими нуклеотидами гиперполяризационно-активируемые каналы ( англ. Hyperpolarization-activated cyclic nucleotide-gated (HCN) channels ) — это интегральные белки , которые являются лиганд-зависимыми катионными каналами в мембранах клеток сердца и головного мозга . HCN-каналы иногда обозначают как «каналы-водители ритма», поскольку они участвуют в генерации ритмической активности клетками сердца и головного мозга. HCN-каналы кодируются четырьмя генами ( , , , ) и экспрессируются в сердце, центральной и периферической нервной системе и фоторецепторах сетчатки .
Ионные токи через HCN-каналы, обозначаемые как I f ( англ. funny — забавный/странный) или I h ( англ. hyperpolarizing — гиперполяризирующий) в сердце и I h , I q ( англ. query — странный) в нервной системе , играют ключевую роль в контроле над сердечной и нейрональной ритмической активностью («токи-водители ритма»). Такое обозначение (funny и query) они получили из-за того, что их активация происходит при гиперполяризации , в то время как обычно ионные каналы активируются при . Процесс активации канала облегчается при присоединении цАМФ . HCN-каналы не инактивируются при деполяризации. В нервной системе они, по всей видимости, отвечают за регуляцию цикла сон-бодорствование, ритм дыхания и ритмической активности, участвующей в синхронизации различных областей мозга, необходимой для их совместной работы .
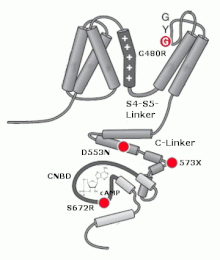
HCN-каналы принадлежат к суперсемействам (Kv) и каналов, управляемых циклическими нуклеотидами (CNG), что отличает их от остальных потенциал-зависимых каналов. HCN-каналы состоят из 4 субъединиц, которые могут быть как одинаковыми, так и отличаться друг от друга . Однако, in vivo чаще встречаются каналы, состоящие из субъединиц одного типа . Каждая субъединица содержит шесть трансмембранных (S1—6) доменов, включая позитивно заряженный потенциалзависимый сенсор (S4), область поры, расположенную между S5 и S6 и несущую GYG мотив калиевых каналов и домен связывания циклических нуклеотидов (CNBD) на С-конце. Изоформы HCN высококонсервативны относительно трансмембранных доменов и участка связывания циклических нуклеотидов (на 80—90 % идентичны), но различаются своими амино- и карбокси-концами . HCN-каналы, состоящие из субъединиц одного типа, отличаются по своим свойствам. Так, наиболее чувствительными к регуляции через цАМФ являются HCN2 и HCN4. Наибольшей скоростью активации обладают HCN1-каналы, а наименьшей HCN4 .
В синоатриальном узле наиболее распространённой является изоформа HCN4, однако, изоформы HCN1 и HCN2 так же встречаются, хоть и в гораздо меньших количествах. Ионный ток через HCN каналы, называемый «funny» (забавный) или пейсмейкерным током (I f ), играет ключевую роль в генерации и регуляции .
Все четыре типа субъединиц HCN-канала экспрессируются в мозге . HCN1 встречается чаще всего в коре головного мозга во всех слоях , гиппокампе , коре мозжечка и стволе мозга . HCN2 экспрессируется в таламусе , бледном шаре , ядрах ствола мозга. HCN3 характерен для нижних отделов нервной системы. HCN4 присутствует в ядрах таламуса, базальных ганглиях и . Обнаруживаются HCN-каналы и в периферической нервной системе . В дополнение к их роли в качестве водителей ритмической или осцилляторной активности, HCN-каналы могут регулировать возбудимость нейронов. Так, в пирамидальных нейронах коры и гиппокампа HCN-каналы расположены, в основном, на апикальных дендритах , регулируя их возбудимость и связанность нейронной сети . Некоторые исследование позволяют судить об их роли в восприятии кислого вкуса , координации моторных актов и некоторых аспектов обучения и памяти . Существуют доказательства, что HCN-каналы играют определённую роль в развитии эпилепсии и невропатической боли . Было показано, что HCN-каналы участвуют в сенсорных нейронов обонятельного анализатора, определяемого их активностью .

















































