Interested Article - Брахиозавр

- 2021-04-04
- 1
Брахиоза́вр ( лат. Brachiosaurus — плечистый ящер, от др.-греч. βραχίων [ brachion ] — плечо и др.-греч. σαῦρος [ sauros ] — ящер, ящерица) — род завроподовых динозавров , живших на территории современной Северной Америки в конце юрского периода , около 154–153 млн лет назад. Включает единственный вид — Brachiosaurus altithorax ( лат. altithorax — глубокогрудый). Описан в 1903 году американским палеонтологом Элмером Риггсом по ископаемым остаткам, обнаруженным в долине реки Колорадо в западной части Колорадо , США . Прижизненная длина известных образцов брахиозавра, по разным оценкам, составляла от 18 до 21 метра; оценки массы варьируются от 28,3 до 58 тонн . У брахиозавра была непропорционально длинная шея и небольшой череп, что в целом типично для завропод. Передние конечности были длиннее задних, в результате чего туловище имело весьма крутой наклон, а хвост казался пропорционально коротким .
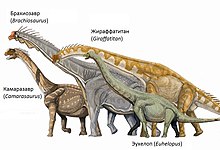
Брахиозавр — типовой род семейства брахиозаврид (Brachiosauridae). В значительной степени представления о брахиозавре основываются на данных об известном по более полному материалу брахиозавриде жираффатитане , описанному из в Танзании . Жираффатитан был выделен в 1914 году немецким палеонтологом Вернером Яненшом как вид брахиозавра ( B. brancai ), но перенесён в собственный род в 2009 году. Три других вида брахиозавра были описаны по остаткам, найденным в Африке и Европе; два из них более не считаются валидными или диагностируемыми, а третий выделен в собственный род .
Голотип B. altithorax (экземпляр FMNH P 25107) является наиболее полным известным науке образцом вида. Известно лишь несколько других образцов, что делает брахиозавра одним из самых редких завропод . Брахиозавр считается растительноядным животным, обрезавшим или щипавшим листья деревьев на высоте до 9 м от земли . В отличие от ряда других завропод, он вряд ли был способен с лёгкостью подниматься на задние конечности . Некоторые учёные приводили брахиозавра в качестве примера динозавра, который, скорее всего, был эктотермным («холоднокровным») из-за его большого размера и соответствующей потребности в достаточном количестве пищи, но современные исследования показывают, что он, как, вероятно, и другие динозавры, был , либо (« теплокровным ») . Как один из наиболее знаковых и первоначально считавшийся одним из крупнейших динозавров, брахиозавр обрёл известность в популярной культуре , в частности, он появляется в фильме « Парк юрского периода » (1993) .
История открытия
Голотип

Род Brachiosaurus описан по фрагментарному посткраниальному скелету, обнаруженному в 1900 году в долине реки Колорадо близ города в Колорадо . Этот скелет, который позже будет выбран в качестве голотипа , происходит из пород бассейна Браши-Мембер формации Моррисон и, следовательно, является позднекиммерийским по возрасту; возраст отложений составляет примерно 154–153 млн лет . Скелет был обнаружен американским палеонтологом Элмером Риггсом и его коллегами из Колумбийского музея Филда (теперь Филдовский музей естественной истории ) в Чикаго , и получил каталожный номер FMNH P 25107 .
Риггс и коллеги начали раскопки в результате благоприятной переписки со Стэнтоном Мериллом Брэдбери, дантистом из близлежащего Гранд-Джанкшена . Весной 1899 года Риггс разослал мэрам западного Колорадо письма, в которых интересовался возможными тропами, ведущими от железнодорожных станций на северо-восток Юты, где он надеялся найти окаменелости эоценовых млекопитающих . К удивлению Риггса, Брэдбери, который был коллекционером-любителем и президентом Академии наук Западного Колорадо, сообщил ему, что с 1885 года недалеко от Гранд-Джанкшн неоднократно находили кости динозавров . Риггс скептически отнесся к этому заявлению, но его начальник, куратор геологии Оливер Каммингс Фаррингтон, хотел добавить в коллекцию большой скелет завропода, чтобы превзойти другие учреждения, и убедил руководство музея вложить пятьсот долларов в экспедицию .
Прибыв 20 июня 1900 года, палеонтологи разбили лагерь на заброшенном ранчо Гот-Ранч ( англ. Goat Ranch) . Во время разведывательной поездки верхом на лошади полевой помощник Риггса Гарольд Уильям Менке обнаружил плечевую кость FMNH P 25107 ; 4 июля он воскликнул, что это «самая большая вещь на свете!» ( англ. the biggest thing yet!). Сначала Риггс принял находку за плохо сохранившийся образец апатозавра или бронтозавра и отдал приоритет раскопкам в карьере №13 ( англ. Quarry 13), в котором находился более многообещающий скелет (теперь считается синонимом камаразавра ). Удостоверившись в наличии скелета, 26 июля он вернулся к плечевой кости в 13-м карьере, которая, как вскоре выяснилось, оказалась действительно огромных размеров, убедив Риггса в том, что он обнаружил самое большое наземное животное в истории . В 1904 году Риггс отметил: «Если бы не необычный размер рёбер , связанных с ним, образец был бы отброшен как апатозавр, слишком плохо сохранившийся, чтобы представлять ценность» ( англ. Had it not been for the unusual size of the ribs found associated with it, the specimen would have been discarded as an Apatosaur, too poorly preserved to be of value) .

Место обнаружения голотипа, Riggs Quarry 13 , расположено на небольшом холме, ныне известном как холм Риггса ( англ. Riggs Hill); в настоящее время оно отмечено мемориальной доской. Сообщалось о ещё нескольких окаменелостях брахиозавров, обнаруженных на холме Риггса, но эти находки подверглись вандализму . Во время раскопок голотипа Риггс ошибочно идентифицировал плечевую кость как деформированную бедренную кость из-за её большой длины, и это, казалось, подтвердилось, когда была обнаружена хорошо сохранившаяся настоящая бедренная кость такого же размера от того же скелета. Только после препарации ископаемого материала в лаборатории кость была признана плечевой . Раскопки привлекли большое количество посетителей, что задержало ход работы и вынудило Менке охранять участок для предотвращения кражи костей. 17 августа последняя кость была покрыта гипсом . После заключительной десятидневной исследовательской поездки экспедиция вернулась в Гранд-Джанкшен и наняла команду и фургон для перевозки всех окаменелостей на железнодорожную станцию, что заняло пять дней; ещё одна неделя ушла на то, чтобы упаковать находки в тридцать восемь ящиков массой примерно в 5 700 кг. 10 сентября Риггс уехал поездом в Чикаго, прибыв туда 15-го числа; железнодорожная компания разрешила перевозку пассажиров и грузов в качестве пиар-жеста .
Скелет голотипа состоит из правой плечевой кости, правой бедренной кости, правой подвздошной кости , правого коракоида , крестца , последних семи туловищных позвонков, двух хвостовых позвонков и нескольких рёбер . Риггс описал коракоид как находившийся на левой стороне тела , но повторное исследование показало, что это был правый коракоид . Нижний конец плечевой кости, нижняя часть крестца, подвздошная кость и сохранившиеся хвостовые позвонки в той или иной степени пострадали от выветривания. Позвонки оказались лишь немного смещены из своего первоначального анатомического положения; их обнаружили так, что они были направлены верхней частью вниз. Однако рёбра, плечевая кость и коракоид были смещены влево от позвоночника, что указывает на то, что кости переносились водным потоком. Об этом также свидетельствует изолированная подвздошная кость диплодока , которая, по-видимому, дрейфовала против позвоночника брахиозавра, а также изменения в составе окружающих пород. Хотя образец сохранился, по большей части, в тонком слое мелкозернистой глины, у седьмого предкрестцового позвонка этот слой заменяется толстым слоем гораздо более крупных отложений, состоящих из гальки у их основания и песчаника выше, что говорит об образовании отложений в мощном потоке. Основываясь на этих доказательствах, Риггс в 1904 году предположил, что недостающую переднюю часть скелета смыл поток воды, тогда как задняя уже была покрыта отложениями, благодаря чему и сохранилась .
Риггс опубликовал краткий отчёт о находке в 1901 году, отметив необычную длину плечевой кости относительно бедренной, чрезвычайно большой общий размер тела и вытекающие отсюда жирафоподобные пропорции, а также меньшее развитие хвоста, но не дал новому динозавру научного названия . В 1903 году он описал род Brachiosaurus с типовым и единственным видом Brachiosaurus altithorax . Родовое название относится к необычной для завропод длине передних конечностей: оно происходит от др.-греч. βραχίων [ brachion ] — «плечо» и др.-греч. σαῦρος [ sauros ] — «ящер, ящерица», означая, таким образом, «плечистый ящер». Видовое название было выбрано из-за необычайно глубокой и широкой грудной полости : оно произведено от лат. altus — «глубокий» и др.-греч. θώραξ [ thorax ] — «нагрудник, кираса , корслет ». Латинское thorax пришло из древнегреческого и стало научным названием грудной клетки . В названиях статей Риггса 1901 и 1903 годов подчёркивалось, что брахиозавр — «крупнейший из известных динозавров» ( англ. largest known dinosaur) . За публикацией 1903 года последовало более подробное описание брахиозавра в монографии Риггса 1904 года .

Препарация голотипа началась осенью 1900 года, вскоре после того, как Риггс отвёз его в Филдовский музей естественной истории . Сначала препарировались элементы конечностей. Зимой 1904 года сильно выветренные спинные позвонки и бедренная кость были отпрепарированы Джеймсом Б. Эбботтом и К. Т. Клайном . Когда препарация каждой кости была завершена, находки выставили в витрине зала 35 Дворца изящных искусств на Всемирной выставке 1893 года , первого месторасположения Филдовского музея. Все кости всё ещё экспонировались по отдельности в зале 35 к 1908 году, когда публике был представлен недавно смонтированный «апатозавр» FMNH P25112 (позже переклассифицирован как бронтозавр ), остатки которого Риггс обнаружил в карьере 12 . Попытки смонтировать скелет брахиозавра не предпринималось, потому что удалось обнаружить только 20% скелета. В 1993 году были сделаны слепки с костей голотипа, а недостающие кости вылепили на основе материала родственного Brachiosaurus brancai (ныне Giraffatitan ) из Музея естествознания в Берлине . Этот пластиковый скелет был установлен и в 1994 году выставлен на всеобщее обозрение в северной части зала Стэнли Филда, главного выставочного зала нынешнего здания Филдовского музея. Настоящие кости голотипа выставлялись в двух больших стеклянных витринах по обеим сторонам от слепка. Слепок простоял до 1999 года, когда его переместили в зал B терминала United Airlines в Международном аэропорту О’Хара , чтобы освободить место для недавно приобретенного музеем скелета тираннозавра по прозвищу . В то же время Филдовский музей установил второй пластиковый слепок скелета, предназначенный для использования вне помещений, который выставили за пределами музея на северо-западной террасе . Ещё один «уличный» слепок был отправлен в , где теперь он служит частью ворот-моста в «DinoLand, U.S.A.» .
Дальнейшие открытия
Дальнейшие открытия окаменелостей брахиозавра в Северной Америке были редки, причём новые находки представлены только несколькими костями. Поскольку у голотипа не сохранились череп, шея, передние спинные позвонки, задние конечности и ступни, ряд других образцов, потенциально принадлежащих брахиозавру, невозможно сравнить с голотипом и, соответственно, уверенно установить их видовую принадлежность. Тем не менее, материал был описан из Колорадо , Оклахомы , Юты и Вайоминга , а также упоминался неописанный материал из некоторых других регионов .
В 1883 году фермер Маршалл Паркер Фелч, работавший на палеонтолога Отниела Чарлза Марша , сообщил об обнаружении черепа завропода в названном в его честь карьере Фелча ( Felch Quarry 1 ), который расположен недалеко от , Колорадо. Череп был найден в желтовато-белом песчанике, рядом с шейным позвонком длиной в 99 см, который разрушился при попытке извлечь его. Фелч отправил череп Маршу, в , где образец получил каталожный номер YPM 1986. В 1891 году Марш включил череп в свою реконструкцию скелета бронтозавра, возможно, из-за того, что Фелч идентифицировал его как принадлежащий этому динозавру. Череп из карьера Фелча состоит из собственно черепа ( лат. cranium), верхней челюсти , правой задней части челюсти, верхней левой части челюсти, левой чешуйчатой кости , зубных костей и, возможно, неполной крыловидной кости . Кости препарировались достаточно грубо, что привело к их повреждению. Большинство образцов, обнаруженных Фелчем, были отправлены в Национальный музей естественной истории после смерти Марша, в 1899 году, в том числе и череп, которому присвоили новый каталожный номер USNM 5730 .
В 1975 году Джек Макинтош и Дэвид Берман пришли к выводу, что если бы Марш не изобразил на своей скелетной реконструкции бронтозавра с неправильным черепом, по пропорциям схожим с таковым у камаразавра, то « диплодоцидность » черепа первого могла бы быть установлена несколько ранее. Макинтош и Берман отметили, что разрушенный позвонок, найденный Фелчем, мог принадлежать брахиозавру . Позже Макинтош предположил, что череп из карьера Фелча, вероятно, также принадлежит брахиозавру, и обратил на него внимание палеонтологов Кеннета Карпентера и Вирджинии Тидвелл, призвав их описать его. Они привезли череп в , где лучше отпрепарировали его и сделали реконструкцию на основе слепков отдельных костей, при этом черепа жираффатитана и камаразавра послужили образцами при восстановлении недостающих костей .

В 1998 году Карпентер и Тидвелл описали череп из карьера Фелча и официально классифицировали его как относящийся к брахиозавру неопределённого вида ( Brachiosaurus sp.). Авторы обосновали отнесение черепа к брахиозавру его сходством с черепом B. brancai , теперь известного как Giraffatitan ; при этом они не причислили череп непосредственно к виду B. altithorax из-за отсутствия перекрывающегося материала . В 2020 году было формально опубликовано исследование Майкла Д. Д'Эмика и Мэтью Т. Каррано, в котором они переописали череп из карьера Фелч после проведения его дальнейшей препарации и компьютерной томографии , а также сверки с историческими иллюстрациями. Палеонтологи пришли к выводу, что квадратная кость и зуб из зубной кости, которые Карпентер и Тидвелл считали частью черепа, на самом деле не относятся к нему. Квадратная кость слишком велика, чтобы она могла быть сочленена с чешуйчатой, сохранилась иначе, чем другие кости, и была обнаружена в нескольких метрах от них. Зуб не похож на зубы внутри челюстей (по данным компьютерной томографии), он превышает их по размеру и поэтому был отнесен к Camarasaurus sp. (из карьера известны и другие зубы, относимые к этому роду). Кроме того, Д'Эмик и Каррано сочли наиболее экономным отнести череп к самому B. altithorax , а не к неустановленному виду, поскольку нет каких-либо доказательств наличия других таксонов брахиозаврид в формации Моррисон, а включение этого и другого возможного материала брахиозавра в филогенетический анализ не повлияло на положение B. altithorax .
Виды

Хотя в настоящее время Brachiosaurus altithorax обычно считается единственным известным видом брахиозавров , ранее к роду относили несколько других видов. Между 1909 и 1912 годами в ходе серии крупномасштабных Германских Восточно-Африканских палеонтологических экспедиций было обнаружено значительное количество материала брахиозаврид из формации Тендагуру. В 1914 году немецкий палеонтолог Вернер Яненш сравнил находки с окаменелостями B. altithorax и сделал вывод, что все они могут быть объединены в род Brachiosaurus . По этому материалу Яненш описал два вида: крупный и представленный почти полным скелетом Brachiosaurus brancai и более мелкий фрагментарный Brachiosaurus fraasi . В трёх последующих публикациях 1929 , 1950 и 1961 гг. Яненш сравнил виды более подробно, обнаружив тринадцать общих признаков, характерный как для Brachiosaurus brancai (с которым он синонимизировал B. fraasi ) и B. altithorax . В публикации Майкла Тайлора 2009 года за действительные принимаются только четыре из этих признаков; шесть отнесены к группам, более инклюзивным , чем брахиозавриды, один на самом деле отсутствует у B. brancai , а действительность ещё двух затруднительно оценить .
В 1957 году и Жорж Збышевски описали из отложений титонского яруса верхней юры Аталайе в Португалии вид Brachiosaurus atalaiensis . В 2003 году Мигел Теллеш Антунеш и Октавио Матеуш переместили этот вид в собственный род Lusotitan , отнесённый к семейству брахиозаврид .
был выделен де Лаппарентом в 1960 году ; определён как новый вид брахиозавра, описанный по части крестца, четырём крестцовым позвонкам и фрагментов передней конечности, найденных в местности Wargla в Алжире . Позже было установлено, что данные окаменелости относятся к альбскому веку , и примерно на 40 миллионов лет моложе Brachiosaurus altithorax и Giraffatitan . В настоящее время таксон помечен как nomen dubium , вероятно, из-за низкого качества ископаемого материала . Кости были обнаружены далеко от друга, поэтому Маннион и коллеги (2013) отметили, что они могут принадлежать разным видам .
-
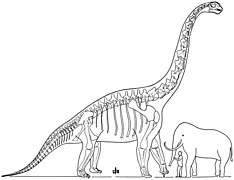 Диаграмма, включающая кости как брахиозавра, так и жираффатитана , за авторством Уильяма Диллера Мэтью , 1915
Диаграмма, включающая кости как брахиозавра, так и жираффатитана , за авторством Уильяма Диллера Мэтью , 1915 -
![Средние хвостовые позвонки Lusotitan[en], ранее известного как "B." atalaiensis](/images/001/073/1073788/12.jpg?rand=719323) Средние хвостовые позвонки , ранее известного как "B." atalaiensis
Средние хвостовые позвонки , ранее известного как "B." atalaiensis -
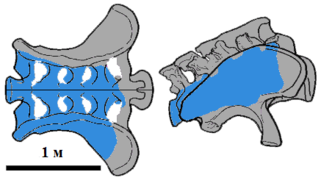 Крестец "B." nougaredi ; известные элементы отмечены голубым цветом
Крестец "B." nougaredi ; известные элементы отмечены голубым цветом
Описание
Размер

Большинство оценок размера Brachiosaurus altithorax основаны на данных о родственном ему брахиозавриде жираффатитане (ранее B. brancai ), который известен по гораздо более полному материалу, чем брахиозавр. Брахиозавр и жираффатитан являются самыми крупными брахиозавридами, которым принадлежат относительно полные ископаемые остатки. Следует учитывать, что типовой и наиболее полный образец брахиозавра, по-видимому, принадлежал молодой особи, о чём свидетельствуют несочленённый между коракоидом, костью плечевого пояса , образующей часть плечевого сустава , а также строение лопатки . На протяжении многих лет были даны разные оценки массы B. altithorax : 28,3 т , 28,7 т , 35 т , 40 т , 43,9 т , 56,3 т и 58 т . Длина брахиозавра оценивалась в 18 м , 20–21 м и 24,5 м , а его высота — в 9,4 м и 12–13 м .
Из материала, возможно, принадлежащего ещё более крупному брахиозавру, известна гигантская неполная бедренная кость Brachiosaurus sp. из Юты , США; из-за неполноты кости не исключается также возможность её атрибуции другому роду (например, камаразавру ). Р. Молина-Перес и А. Ларраменди (2020) оценили длину животного, которому принадлежала кость, в 26,5 м, а его массу — в 50 т .
Общий план строения

Как и другие завроподы, брахиозавр был квадропедальным («четвероногим») динозавром с маленьким черепом, длинной шеей, большим туловищем, длинным мускулистым хвостом и тонкими столбообразными конечностями . Многочисленные воздушные мешки , связанные с дыхательной системой , присутствовали в шее и туловище, врезаясь в позвонки и рёбра путём костной резорбции , при этом значительно снижая общую плотность тела . У голотипа не сохранилась шея, но она, скорее всего, была очень длинной даже по стандартам завропод, о чём можно судить по остаткам близкородственного брахиозавру жираффатитана, шея которого состояла из тридцати удлинённых шейных позвонков . Шея держалась в небольшом S-образном изгибе, в котором нижняя и верхняя части шеи были согнуты, а средняя оставалась прямой .
Как и у жираффатитана, у брахиозавра, вероятно, имелись сильно удлинённые , которые опускались до нижней части шеи, перекрывая несколько предшествовавших позвонков. Окончания таких удлинённых рёбер прикреплялись к шейным мышцам, располагавшимся ближе к туловищу, что позволяло этим мышцам управлять удалёнными областями шеи, при этом не перегружая их лишним весом . По всей видимости, брахиозавр и жираффатитан имели небольшой плечевой горб между третьим и пятым спинными позвонками, в котором боковые и направленные вверх позвонков были длиннее, обеспечивая дополнительную поверхность для прикрепления шейных мышц .
Грудная клетка была глубокой по сравнению с такой у других завропод . Хотя плечевая и бедренная кости были примерно равны по длине, вся передняя конечность обладала большей длиной в сравнении с задней, так как у других брахиозаврид имелись удлинённые предплечье и пястная кость . В результате того, что передние конечности превышали по длине задние, туловище держалось в наклоненной позиции, при которой плечо располагалось намного выше бёдер, а шея выходила из туловища под крутым углом. Таким образом, по своему строению брахиозавр в общих чертах напоминал жирафа . При этом, у большинства других завропод передняя конечность короче задней; передняя конечность была особенно коротка у диплодокоид , живших в одно время с брахиозавром .
Брахиозавр отличался по пропорциям тела от близкородственного жираффатитана. Длина туловища составляла примерно 25–30 % от всей длины позвоночника, и спинная часть позвоночника была длиннее плечевой кости. Обнаружен только один полный хвостовой позвонок , но его высота позволяет предположить, что хвост был больше, чем у жираффатитана. Этот позвонок имел гораздо бо́льшую площадь для прикрепления связки благодаря расширенным невральным отросткам , что также указывает на то, что хвост был также длиннее, чем у жираффатитана, возможно, на 20–25 % . В 1988 году палеонтолог Грегори Пол предположил, что шея брахиозавра была короче, чем у жираффатитана, но в 2009 году другой палеонтолог, Майк Тайлор, указал, что два шейных позвонка, вероятно, принадлежащие брахиозавру, имеют одинаковые пропорции . В отличие от жираффатитана и других завропод, у которых имелись вертикальные столбообразные передние конечности, плечи брахиозавра, по-видимому, были слегка растянуты в плечевых суставах, на что указывает боковая ориентация суставных поверхностей коракоидов. Плечевая кость была менее стройной, чем у жираффатитана, в то время как бедренная кость имела аналогичные пропорции. Это может указывать на то, что передние конечности брахиозавра поддерживали бо́льшую массу тела, чем конечности жираффатитана .
Посткраниальный скелет

Известна только часть спинных позвонков брахиозавра; в полностью сохранившемся позвоночнике образца брахиозаврида BMNH R5937, видовая принадлежность которого пока не определена, имеется 12 спинных позвонков . Передние спинные позвонки брахиозавра несколько выше, но намного длиннее, чем задние. Данная особенность является одной из черт, отличающих брахиозавра от жираффатитана, у которого передние спинные позвонки, в сравнении с задними, значительно выше, но лишь немного длиннее. У брахиозавра тела позвонков более вытянутые и относительно круглые в поперечном сечении, в то время как у жираффатитана они больше в широту, чем в высоту. Небольшие отверстия ( лат. foramina) по бокам тел позвонков брахиозавра, через которые внутрь проникали воздушные мешки, больше, чем у жираффатитана. Диапофизы брахиозавра горизонтальные, а у жираффатитана наклонены вверх. На своих концах диапофизы сочленялись с рёбрами; тогда как у жираффатитана суставная поверхность отчётливо треугольная, у брахиозавра «треугольность» не столько выражена. При виде сбоку невральные отростки позвонка брахиозавра стоят вертикально и в два раза шире в основании, чем на вершине; у жираффатитана же они отклоняются назад и не расширяются в своей основе. При взгляде спереди или сзади, видно, что невральные отростки как брахиозавра, так и жираффатитана, расширяются кверху .
В случае с брахиозавром невральные отростки расширяются кверху постепенно, в результате чего они приобретают веслоподобную форму; у жираффатитана расширение появляется внезапно и только в самой верхней части отростка. Как на передней, так и на задней сторонах невральных отростков имелись большие треугольные и бугристые поверхности, которые у жираффатитана, в отличие от брахиозавра, полукруглые и намного меньше. Различные отростки позвонков соединялись несколькими типами листоподобных или похожих на выпирающие гребни костных пластинок ( лат. laminae), для которых выработана особая номенклатура. У брахиозавра отсутствуют postspinal laminae , но у жираффатитана они проходят по задней стороне неврального отростка. От невральных отростков брахиозавра к диапофизам проходят так называемые spinodiapophyseal laminae , объединённые с spinopostzygapophyseal laminae , которые проходят между невральными и в задних частях позвонков, доходя до середины невральных отростков. У жираффатитана обе этих пластинки не объединены, и spinodiapophyseal laminae доходят до верхушек невральных отростков. Брахиозавр отличается от жираффатитана отсутствием трёх деталей в пластинках спинных позвонков, которые являются уникальными для последнего рода .

Воздушные мешки проникали не только в позвонки, но и в рёбра. У брахиозавра проникновение воздушных мешков осуществлялось через небольшое отверстие на передней стороне рёбер, тогда как у жираффатитана отверстия присутствуют и передней, и на задней сторонах бугорка ( лат. tuberculum), костного образования, сочленённого с диапофизами. Пол в 1988 году заявил, что рёбра брахиозавра были длиннее, чем у жираффатитана, но это утверждение было поставлено под вопрос Тейлором в 2009 году . За спинной частью позвоночника располагается крестец , состоящий из пяти окостеневших вместе крестцовых позвонков . Как и у жираффатитана, крестец брахиозавра пропорционально широкий и имеет очень короткие невральные отростки. Плохая сохранность материала крестца жираффатитана препятствует детальному сравнению обоих родов по особенностям его анатомии .
От хвоста брахиозавра из известного материала хорошо сохранился только второй хвостовой позвонок. Как и у жираффатитана, этот позвонок слегка амфицельный (вогнутый с обоих концов), без отверстий по бокам и с коротким прямоугольным невральным отростком, наклонённым назад. В отличие от второго хвостового позвонка у жираффатитана, у брахиозавра на этом позвонке имеется пропорционально более высокая невральная дуга, что делает его примерно на 30 % выше. На хвостовом позвонке брахиозавра отсутствуют вдавливания по бокам, имеющиеся у жираффатитана. При виде спереди или сзади невральный отросток хвостового позвонка брахиозавра расширяется к своей вершине так, что его ширина примерно в три раза превышает минимальную, но у жираффатитана не наблюдается никакого расширения. Невральный отросток хвостового позвонка брахиозавра отклоняется назад примерно на 30°, а у жираффатитана — на 20°. Хвостовые рёбра выступают в бок и не отклоняются назад, как у жираффатитана. Суставные грани суставных отростков в задней части позвонка брахиозавра направлены вниз, в то время как у жираффатитана они в большей степени обращены в сторону. Помимо того, что сочленение между позвонками формировалось суставными отростками, также присутствовало дополнительное , делавшее позвоночник более жёстким; у брахиозавра гипосфен гораздо более выражен, чем у жираффатитана .
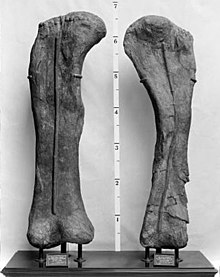
Коракоид полукруглый и более высокий, чем широкий; при виде сбоку наблюдаются отличия коракоида брахиозавра от такого у жираффатитана. Шов на лопатке брахиозавра более прямой. Кроме того, суставная поверхность, которая образует часть плечевого сустава, толще и более выраженнее в сторону, чем у жираффатитана и других завропод, что, возможно, указывает на более широко расставленные конечности брахиозавра. Сохранившиеся элементы плечевой кости достигают 204 см в длину, хотя часть её нижнего конца была утеряна из-за эрозии; полная длина кости оценивается в 216 см. У брахиозавра плечевая кость более тонкая, чем у большинства других завропод: её ширина составляет всего 28,5 см в самой узкой части. При этом она была более прочной, чем у жираффатитана, и примерно на 10 % шире в верхних и нижних концах. На верхнем конце она имеет низкую выпуклость, видимую сбоку, которая отсутствует у жираффатитана .
Отличительные признаки также обнаружены в подвздошной кости таза. У брахиозавра седалищая ножка, нисходящая вниз и соединяющаяся с седалищной костью , простирается ещё ниже, чем у жираффатитана. У представителей последнего рода имелась резкая выемка между седалищной ножкой и задней частью подвздошной кости, а у брахиозавра эта выемка более округлая. На верхней поверхности задней части подвздошной кости брахиозавра находится выраженный бугорок, которого нет у других завропод. Бедренная кость брахиозавра очень похожа на такую у жираффатитана, но немного более крепкая и была измерена как достигающая 203 см в длину . Как и у жираффатитана, по форме она сильно эллиптическая в поперечном сечении и более чем в два раза шире при виде спереди или сзади, чем при виде сбоку. Четвёртый вертел , заметная выпуклость на задней поверхности бедренной кости, более заметен и расположен ниже, чем у жираффатитана. Эта выпуклость служила опорной точкой для одной из наиболее важных двигательных мышц всех длиннохвостых динозавров, хвостово-бедренной мышцы , находившейся в хвосте и оттягивавшей верхнюю часть бедра назад при сжатии. На нижнем конце бедренной кости пара мыщелков не простирается назад так сильно, как у жираффатитана; оба мыщелка схожи по ширине у брахиозавра, но различаются у жираффатитана .
Череп
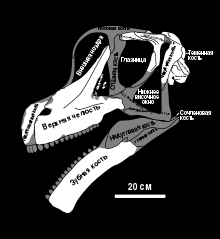
Известный материал
Неизвестный материал
По реконструкции Карпентера и Тидуэлла (1998), череп брахиозавра, обнаруженный в карьере Фелча, достигал около 81 см в длину от в задней части черепа до передней части предчелюстной кости , что делает его самым большим черепом завропода из формации Моррисон . Однако Де Эмик и Каррано (2020) дали другую оценку, по которой череп достигал 70 см в длину и, если он был пропорционален черепу жираффатитана, около 55 см в высоту и 35 см в ширину . В целом, череп высокий, как и у жираффатитана, с длинной мордой (около 36 % длины черепа согласно Карпентеру и Тидуэллу) перед типичной для брахиозаврид перегородкой между ноздрями. Морда притупленная при виде сверху (как у жираффатитана) и, поскольку она расположена под углом относительно остальной части черепа, создаётся впечатление, что она направлена вниз .
Верхнее и боковое височные окна большие, возможно, из-за силы, передаваемой им от массивной челюстной приводящей мускулатуры. Лобные кости на верхней части черепа короткие и широкие, как и у жираффатитана, сросшиеся и соединённые швом с теменными костями , которые сами сочленены вместе. Поверхность теменных костей между верхними височными окнами шире, чем у жираффатитана, но уже, чем у камаразавра . Череп брахиозавра отличается от черепа жираффатитана U-образным швом между лобной и носовой костями (у второго он W-образный), форма которого более выражена благодаря лобным костям, проходящим вперёд над глазницами . Как и у жираффатитана, шейка затылочного мыщелка брахиозавра очень длинная .
Палеобиология
Среда обитания
На протяжении XIX и начала XX веков считалось, что завроподы, такие как брахиозавр, были слишком массивными, чтобы выдерживать собственную массу на суше, и поэтому проводили всю свою жизнь в воде . Риггс, подтверждая наблюдения Джона Белла Хэтчера , был первым, кто в своём отчете 1904 года о брахиозаврах предпринял попытку отстоять гипотезу о том, что большинство завропод были полностью наземными животными, указав, что их полые позвонки не имеют аналогов у современных водных или полуводных животных, а длинные конечности и компактные ступни этих динозавров указывают на их наземную локомоцию. Адаптированность брахиозавра к передвижению по суше оказалась особенно выражена из-за его тонких конечностей, высокой грудной клетки, широких бедренных костей, высокой подвздошной кости и короткого хвоста. Зигапофизы на спинных позвонках брахиозавра сильно уменьшены, в то время как гипосфен-гипантральное сочленение чрезвычайно развито, в результате чего туловище было жёстким, неспособным сгибаться в стороны. Тело было приспособлено только для передвижения по суше на четырёх конечностях . Хотя идеи Риггса постепенно были забыты в течение первой половины XX века, представление о завроподах как о наземных животных стало получать поддержку с 1950-х годов и теперь повсеместно признано палеонтологами . По мнению палеонтологов Стивена и Сильвии Черкасов (1990), брахиозавр мог лишь иногда входить в воду, чтобы охладиться ( терморегуляция ) .
Функция носовых отверстий

Брахиозавр, как и некоторые другие неозавроподы , обладал большими костными носовыми отверстиями, располагавшимися в верхней части черепа. Традиционно считалось, что мясистые ноздри завропод располагались на макушке головы, примерно в задней части костного носового отверстия. При этом завроподы ошибочно рассматривались как полуводные животные, использовавшие свои большие носовые отверстия в качестве трубок при погружении. Американский палеонтолог Лоуренс Уитмер отверг это мнение в 2001 году, указав, что у всех современных наземных позвоночных внешние мясистые ноздри расположены в передней части костной ноздри. Мясистые ноздри завропод должны были быть расположенными в передней части нариальной ямки ( narial fossa ), углубления, которое шло от костной ноздри к кончику морды .
Черкасы указали, что не существует убедительного способа определить, где были расположены ноздри, если только не будет обнаружена голова с отпечатками кожи. Они предположили, что расширенное носовое отверстие могло освобождать место для ткани, связанной с обонянием животного, которая помогала вынюхивать растительность. Они также отметили, что у современных рептилий наличие выпуклых, увеличенных и приподнятых носовых костей можно соотнести с наличием мясистых рогов и бугристых выступов, и что у брахиозавров и других завропод с большими носовыми костями могли быть декоративные носовые гребни .
Высказывалась также более экзотическая версия о том, что у завропод, в том числе и у брахиозавра, мог быть хобот , которая основывается на расположении нариального отверстия. Фабьен Кнолль и его коллеги оспорили это мнение в отношении диплодока и камаразавра в 2006 году, обнаружив, что отверстие для лицевого нерва в их мозговых коробках было небольшим. Таким образом, лицевой нерв не был увеличен, как у слонов, где он участвует в работе сложной мускулатуры хобота. Кнолль и коллеги отметили, что лицевой нерв у жираффатитана (на тот момент B. brancai ) был больше, но гипотеза о наличии у него хобота подкреплена слишком слабой доказательной базой .
Метаболизм
Подобно другим завроподам, брахиозавр, как это обычно в настоящее время считается, был гомойотермом («теплокровность»), то есть поддерживал стабильную внутреннюю температуру тела, и эндотермом, то есть самостоятельно производил тепло . Согласно другой точке зрения, завроподы и многие другие нептичьи динозавры являлись мезотермами, то есть производили тепло самостоятельно, хотя и не контролировали общую температуру тела в той же мере, как это делает большинство млекопитающих и птиц . В любом случае, завроподы, в том числе и брахиозавр, в той или иной мере могли контролировать температуру своего тела («теплокровность»), самостоятельно производя необходимое тепло .
В 1989 году привёл брахиозавра в качестве примера динозавра, для которого эндотермия маловероятна из-за сочетания большого размера, приводящего к перегреву, и высокой потребности в калориях, которые питали бы эндотермичный организм. Большая носовая дуга была объяснена как приспособление для охлаждения мозга, как поверхность для испарительного охлаждения крови . Сандер (2010) обнаружил, что расчёты Рассела основывались на неверных оценках массы тела и не учитывали существование внутренних охлаждавших тело поверхностей, поскольку на момент исследования не было известно о наличии больших воздушных мешков у завропод. Эти неточности привели к завышенной оценке количества производившегося животным тепла и недооценке его потерь .
В культуре

На первых порах Элмер Риггс пытался ограничить осведомлённость общественности об обнаружении ископаемых остатков брахиозавра. 27 июля 1901 года он читал лекцию для жителей Гранд-Джанкшн, которую проиллюстрировал слайдами, проецировавшимися на импровизированный холст « Волшебным фонарём ». Палеонтолог рассказал общую информацию об эволюции динозавров и методах проведения палеонтологических исследований, но при этом не упомянул, что только что нашел впечатляющий образец . Риггс опасался, что группы из других научных учреждений вскоре узнают об открытии и заберут лучшие из оставшихся окаменелостей. Неделю спустя его хозяин Брэдбери опубликовал в местной газете Grand Junction News статью, в которой объявил о находке одного из самых больших динозавров в истории. 14 августа новость попала в The New York Times . Уже тогда завроподовые динозавры привлекали внимание публики из-за своих гигантских размеров, которые часто преувеличивались сенсационными газетами . Риггс в своих публикациях играл на этом, подчёркивая огромные размеры брахиозавра .
Брахиозавр является одним из самых знаковых динозавров , хотя его наиболее популярные изображения основаны на информации об африканском виде B. brancai , который теперь выделяется в собственный род, названный «жираффатитаном» ( Giraffatitan ) . Астероид главного пояса , 1991 GX 7 , в 1991 году был назван . Брахиозавр фигурирует в фильме 1993 года « Парк юрского периода », где он был воссоздан при помощи компьютерной графики . В то время такие эффекты считалось новаторскими, и зрители хорошо восприняли трепет героев фильма, впервые увидевших «воскрешённого» динозавра . Движения брахиозавра из фильма были основаны на походке жирафа в сочетании с массой слона . Позже в фильме присутствует сцена, для съёмки которой использовались аниматронные голова и шея брахиозавра, взаимодействовавшие с персонажами-людьми . Цифровая модель брахиозавра, использованная в «Парке юрского периода», позже стала отправной точкой для моделей ронто из специального выпуска 1997 года фильма « Звездные войны. Эпизод IV: Новая надежда » .
Примечания
- ↑ (англ.) . Paleofile.com . Дата обращения: 20 июня 2021. 22 октября 2017 года.
- Брахиозавр // Брасос — Веш. — М. : Советская энциклопедия, 1971. — ( Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров ; 1969—1978, т. 4).
- ↑ en . (англ.) // New Mexico Museum of Natural History and Science Bulletin. — Albuquerque, New Mexico: New Mexico Museum of Natural History and Science, 2003. — Vol. 23 . 13 июня 2021 года.
- ↑ , " Brachiosaurus altithorax ", pp. 276—280.
- , "Rearing Giants – kinetic-dynamic modeling of sauropod bipedal and tripodal poses" by H. Mallison, pp. 237—250.
- ↑ , " Brachiosaurus ", pp. —221.
- ↑ Turner C. E., Peterson F. Biostratigraphy of dinosaurs in the Upper Jurassic Morrison Formation of the Western Interior, USA (англ.) // Vertebrate Paleontology in Utah. Miscellaneous Publication 99-1 / In D. D. Gillete (ed.). — Salt Lake City, Utah: en , 1999. — P. 77—114 . — ISBN 978-1-55791-634-1 .
- ↑ Riggs E. S. (англ.) // American Journal of Science : journal. — 1903. — Vol. 15 , iss. 88 . — P. 299—306 . — doi : . — Bibcode : . 17 июня 2019 года. — S2CID .
- ↑ Taylor M. P. (англ.) // Journal of Vertebrate Paleontology : journal. — 2009. — P. 787—806 . — ISSN . — doi : . 22 декабря 2018 года. — S2CID .
- , p. 106.
- , p. 105.
- , p. 108.
- , p. 111.
- ↑ Riggs E. S. (англ.) // Geological Series (Field Columbian Museum). — 1904. — Vol. 2 , iss. 6 . — P. 229—247 . 19 августа 2018 года.
- Chenoweth W. L. (англ.) // Paleontology and Geology of the Dinosaur Triangle / In W. R. Averett (ed.). — Grand Junction, Colorado: Museum of Western Colorado, 1987. — P. 97—100 . — ISBN 9999790410 . 24 июня 2021 года. — LCCN . — OCLC .
- Lohman S. W. (англ.) . — Professional Paper 451. — Reston, Virginia: U.S. Geological Survey , 1965. — P. 1—149. 18 июня 2021 года.
- , p. 117.
- , p. 118—119.
- ↑ Riggs E. S. (англ.) // Science : journal. — 1901. — Vol. 13 , iss. 327 . — P. 549—550 . — ISSN . — doi : . — Bibcode : . — . 24 июня 2021 года. — S2CID .
- Tschopp E., Mateus O., Benson R. B. J. (англ.) // en : journal. — 2015. — Vol. 3 . — P. e857 . — ISSN . — doi : . — . 26 июля 2019 года. — S2CID .
- , p. 243.
- (англ.) , The Field Museum (26 November 1999). 2 марта 2000 года. Дата обращения: 20 июня 2021.
- (англ.) . The Field Museum Photo Archives . Архивировано из 18 марта 2009 года.
- (англ.) . WDW Theme Parks . Архивировано из 7 марта 2019 года.
- ↑ Jensen J. A. (англ.) // en : journal. — 1987. — Vol. 47 , iss. 4 . — P. 592—608 . — ISSN . 24 июня 2021 года. — S2CID .
- Curtice B., Stadtman K., Curtice L. (англ.) // The Continental Jurassic: Transactions of the Continental Jurassic Symposium / In M. Morales (ed.). — Flagstaff, Arizona: Museum of Northern Arizona Bulletin, 1996. — Vol. 60 . — P. 87—95 . — ISBN 9780897341196 . 31 июля 2021 года.
- Curtice B., Stadtman K. The demise of Dystylosaurus edwini and a revision of Supersaurus vivianae (англ.) // Western Association of Vertebrate Paleontologists and Southwest Paleontological Symposium – Proceedings 2001 / In R. D. McCord, D. Boaz (eds.). — Mesa Southwest Museum Bulletin, 2001. — Vol. 8 . — P. 33—40 .
- Bonnan M. F., en . (англ.) // PaleoBios. — 2004. — Vol. 24 , iss. 2 . — P. 12—21 . — ISSN .
- ↑ Carpenter K. , Tidwell V. (англ.) // Modern Geology : journal. — 1998. — Vol. 23 , iss. 1—4 . — P. 69—84 . — ISSN .
- Marsh O. C. (англ.) // American Journal of Science : journal. — 1891. — Vol. s3-41 , iss. 244 . — P. 339—342 . — ISSN . — doi : . — Bibcode : . 24 июня 2021 года. — S2CID .
- ↑ McIntosh J. S., Berman D. S. Description of the palate and lower jaw of the sauropod dinosaur Diplodocus (Reptilia: Saurischia) with remarks on the nature of the skull of Apatosaurus (англ.) // en : journal. — 1975. — Vol. 49 , iss. 1 . — P. 187—199 . — JSTOR .
- Tidwell V. Restoring crushed Jurassic dinosaur skulls for display (англ.) // The Continental Jurassic: Transactions of the Continental Jurassic Symposium / In M. Morales (ed.). — Museum of Northern Arizona Bulletin, 1996. — Vol. 60 .
- ↑ D'Emic M. D., Carrano M. T. (англ.) // en . — 2020. — Vol. 303 , iss. 4 . — ISSN . — doi : . — . 27 ноября 2020 года. — S2CID .
- (англ.) информация на сайте Paleobiology Database . (Дата обращения: 19 июня 2021) .
- Janensch W. (нем.) // Archiv fur Biontologie. — 1914. — Bd. 3 . — S. 81—110 . 19 октября 2021 года.
- Janensch W. Material und Formengehalt der Sauropoden in der Ausbeute der Tendaguru-Expedition (нем.) // Palaeontographica. — 1929. — Bd. 2 (Suppl. 7) . — S. 1—34 .
- Janensch W. Die Wirbelsäule von Brachiosaurus brancai (нем.) // Palaeontographica. — 1950. — Bd. 3 (Suppl. 7) . — S. 27—93 .
- Janensch W. (нем.) // Palaeontographica. — 1961. — Bd. 3 (Suppl. 7) . — S. 177—235 . 31 августа 2021 года.
- de Lapparent A.F., Zbyszewski G. (фр.) // Memoire Service Geologique Portugal. — 1957. — Vol. 2 . — P. 1—63 . 30 января 2022 года.
- ↑ en , en . (англ.) // Comptes Rendus Palevol. — 2003. — Vol. 2 , iss. 1 . — P. 77—95 . — ISSN . — doi : .
- ↑ Mannion P. D., Upchurch P., Barnes R. N., en . (англ.) // en . — 2013. — Vol. 168 , iss. 1 . — P. 98—206 . — ISSN . — doi : . — S2CID .
- de Lapparent A. F. (англ.) = Les dinosauriens du "continental intercalaire" du Sahara central // Memoires de la Societe Geologique de France (Nouvelle Series) / Translated by M. T. Carrano. — Paris, 1960. — Vol. 39 , iss. 1–6 , no. 88 . — P. 1—57 . 17 июня 2021 года.
- ↑ , chpt. 13: "Sauropoda" by P. Upchurch, P. M. Barrett, en , pp. —322.
- ↑ Seebacher F. (англ.) // Journal of Vertebrate Paleontology : journal. — 2001. — 26 March (vol. 21 , no. 1). — P. 51—60 . — ISSN . — doi : . 24 июня 2021 года. — S2CID . — CiteSeer x : .
- ↑ en . (англ.) // Hunteria : journal. — 1988. — Vol. 2 , no. 3 . — ISSN . 19 августа 2018 года. — S2CID .
- ↑ , "Chronology of the largest sauropods throughout history", p. 68.
- Benson R. B. J., Campione N. S .E., Carrano M. T., Mannion P. D., Sullivan C., Upchurch P., Evans D. C. (англ.) // PLOS Biology : journal. — 2014. — Vol. 12 , no. 5 . — P. e1001853 . — doi : . — . 27 мая 2022 года. — PMC . — S2CID .
- Benson R. B. J., Hunt G., Carrano M. T., Campione N., Mannion P. Cope's rule and the adaptive landscape of dinosaur body size evolution (англ.) // en : journal. — 2018. — Vol. 61 , no. 1 . — P. 13—48 . — ISSN . — doi : . — . — PMC .
- , "Appendix: Compilation of published body mass data for a variety of basal sauropodomorphs and sauropods" by N. Klein, K. Remes, C. T. Gee, P. M. Sander, pp. 317—320.
- , p. 40.
- en . Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs (англ.) // en : journal. — en , 2003. — Vol. 29 , no. 2 . — P. 243—255 . — ISSN . — doi : . — JSTOR . — S2CID .
- en . The evolution of vertebral pneumaticity in sauropod dinosaurs (англ.) // Journal of Vertebrate Paleontology : journal. — 2003. — Vol. 23 , no. 2 . — P. 344—357 . — ISSN . — doi : . — JSTOR . — S2CID .
- ↑ Taylor M. P., en . (англ.) // en : journal. — 2013. — Vol. 1 . — P. e36 . — ISSN . — doi : . — Bibcode : . — . 6 июня 2021 года. — PMC . — arXiv : .
- ↑ Christian A., Dzemski G. Reconstruction of the cervical skeleton posture of Brachiosaurus brancai Janensch, 1914 by an analysis of the intervertebral stress along the neck and a comparison with the results of different approaches (англ.) // en : journal. — 2007. — Vol. 10 , no. 1 . — P. 38—49 . — ISSN . — doi : . — S2CID .
- Klein N., Christian A., Sander P. M. (англ.) // Biology Letters : journal. — Royal Society Publishing , 2012. — Vol. 8 , no. 6 . — P. 1032—1035 . — ISSN . — doi : . — . 24 июня 2021 года. — PMC . — S2CID .
- Woodruff D. C. Nuchal ligament reconstructions in diplodocid sauropods support horizontal neck feeding postures (англ.) // en : journal. — Taylor & Francis , 2016. — Vol. 29 , no. 3 . — P. 308—319 . — ISSN . — doi : . — S2CID .
- Carrano M. T. // The sauropods: evolution and paleobiology (англ.) / Christina Curry Rogers, Jeffrey Wilson (eds.). — Oakland, California: University of California Press , 2005. — P. 229—251. — ISBN 9780520246232 . — doi : .
- Migeod F. W. H. British Museum East Africa Expedition: Account of the work done in 1930 (англ.) // Natural History Magazine : journal. — 1931. — Vol. 3 . — P. 87—103 .
- Henderson D. M. (англ.) // Proceedings of the Royal Society of London. Series B: Biological Sciences. — 2004. — Vol. 271 , iss. (Suppl. 4) . — P. S180—S183 . — ISSN . — doi : . — . 28 июля 2021 года. — PMC . — S2CID .
- Jensen J. A. Three new sauropod dinosaurs from the Upper Jurassic of Colorado (англ.) // Great Basin Naturalist : journal. — 1985. — Vol. 45 , iss. 4 . — P. 697—709 . — ISSN . — doi : . — S2CID .
- ↑ , pp. .
- Witmer L. M. Nostril Position in Dinosaurs and Other Vertebrates and Its Significance for Nasal Function (англ.) // Science : journal. — 2001. — Vol. 293 , iss. 5531 . — P. 850—853 . — ISSN . — doi : . — . — S2CID . — CiteSeer x : .
- Knoll F., Galton P. M., López-Antoñanzas R. Paleoneurological evidence against a proboscis in the sauropod dinosaur Diplodocus (англ.) // en : journal. — 2006. — Vol. 39 , iss. 2 . — P. 215–221 . — ISSN . — doi : . — S2CID .
- ↑ Sander P. M., Christian A., Clauss M., Fechner R., Gee C. T. (англ.) // Biological Reviews of the Cambridge Philosophical Society : journal. — 2011. — Vol. 86 , iss. 1 . — P. 117—155 . — ISSN . — doi : . — . 9 марта 2021 года. — PMC . — S2CID .
- ↑ Grady J. M., Enquist B. J., Dettweiler-Robinson E., Wright N. A., Smith F. A. (англ.) // Science : journal. — 2014. — Vol. 344 , iss. 6189 . — P. 1268—1272 . — ISSN . — doi : . — . — S2CID .
- , p. 78.
- , p. 114.
- , p. 115.
- , p. 248.
- , p. 249.
- (англ.) . Jet Propulsion Laboratory . Дата обращения: 18 июня 2021. 24 июля 2020 года.
- Williams G. (англ.) . // IAU Minor Planet Center. Дата обращения: 18 июня 2021. Архивировано из 22 января 2012 года.
- ↑ Shay D., Duncan J. The Making of Jurassic Park (англ.) . — New York: Boxtree Ltd, 1993. — P. 99, 133—135. — 195 p. — ISBN 978-1-85283-774-7 .
- Huls A. . (англ.) , The Atlantic (5 April 2013). 26 мая 2021 года. Дата обращения: 18 июня 2021.
- Britton P. (англ.) // Popular Science . — 1993. — P. 86—91 . — ISSN . 24 июня 2021 года.
- (неопр.) . en . StarWars.com . Дата обращения: 18 июня 2021. Архивировано из 3 октября 2008 года.
Литература
- Brinkman P. D. (англ.) / ed. by J. O. Farlow and T. R. Holtz , editors. — Chicago and London: The University of Chicago Press , 2010. — 345 p. — ISBN 978-0226074726 .
- Czerkas S. J., Czerkas S. A. (англ.) . — Limpsfield: Dragons’ World, 1990. — 247 p. — ISBN 978-0-7924-5606-3 .
- en . (англ.) / ed. by J. O. Farlow and T. R. Holtz , editors. — 2nd ed. — Bloomington, Indiana: Indiana University Press , 2020. — 532 p. — (Life of the Past). — ISBN 9780253051585 .
- en . (англ.) . — Jefferson, N.C., and London: McFarland & Company , 1997. — 345 p. — ISBN 978-0-89950-917-4 .
- (англ.) / ed. by N. Klein, K. Remes, C. T. Gee, and P. M. Sander. — Berkeley: Indiana University Press , 2011. — 331 p. — (Life of the Past). — ISBN 978-0-253-35508-9 .
- (англ.) / ed. by D. B. Weishampel , P. Dodson, and H. Osmolska . — 2nd ed. — Berkeley: University of California Press , 2004. — 880 p. — ISBN 978-0-520-25408-4 .
- Molina-Perez R., Larramendi A. Dinosaur Facts and Figures: The Sauropods and Other Sauropodomorphs (англ.) / Illustrations by A. Atuchin and S. Mazzei. — Princeton, New Jersey: Princeton University Press , 2020. — 272 p. — ISBN 978-0-691-19069-3 .
- en . An Odyssey in Time: Dinosaurs of North America (англ.) . — Minocqua, Wisconsin: NorthWord Press, 1989. — 240 p. — ISBN 978-1-55971-038-1 .
- 2021-04-04
- 1
| † Брахиозавр | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен:
Царство:
Подцарство:
Без ранга:
Без ранга:
Тип:
Подтип:
Инфратип:
Надкласс:
Клада:
Клада:
Клада:
Клада:
Клада:
Клада:
Клада:
Подотряд:
Инфраотряд:
Клада:
Клада:
Семейство:
Род:
†
Брахиозавр
|
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Brachiosaurus Riggs , 1903 | |||||||||||||||||||||||||||||||||||||||||
| Синонимы | |||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||
| Единственный вид | |||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||
|
Геохронология
154—153 млн лет
◄
Наше время
◄
Мел-
палеогеновое вымирание
◄
Триасовое вымирание
◄
Массовое пермское вымирание
◄
Девонское вымирание
◄
Ордовикско-силурийское вымирание
◄
Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||

