Interested Article - Гломеромицеты

- 2020-06-16
- 2
Гломеромице́ты ( лат. Glomeromycetes) — класс грибов , выделяемый в монотипный подотдел Glomeromycotina в составе отдела , содержит около 230 видов ; ранее его представителей рассматривали в составе отдела зигомицетов . Древнейшие ископаемые остатки , достоверно принадлежащие гломеромицетам, имеют возраст 460 млн лет.
Практически все гломеромицеты образуют арбускулярную микоризу с наземными растениями , в том числе с некоторыми мохообразными , однако у них не обнаружено специфичности в отношении растений-хозяев. Не образующие микоризу гломеромицеты обладают внутриклеточными симбионтами — цианобактериями из рода Nostoc . Половой процесс у подавляющего большинства представителей отсутствует. Из-за скрытного образа жизни данных о географическом распространении гломеромицетов практически нет.
Арбускулярная микориза может повышать урожайность сельскохозяйственных культур, таким образом, гломеромицеты могут иметь важное хозяйственное значение.
Строение и биология

Практически все гломеромицеты являются облигатными симбиотрофами и образуют эндомикоризу (арбускулярную микоризу) с более чем 80 % исследованных видов наземных растений . Этот вид микоризы преобладает в саваннах , пустынях и тропических лесах , а в умеренных широтах она встречается у 8 из 10 видов травянистых растений. Гломеромицеты образуют микоризу с такими хозяйственно важными растениями, как злаки . Они образуют микоризу не только со многими видами цветковых растений, но также с некоторыми голосеменными , мохообразными и сосудистыми споровыми растениями .
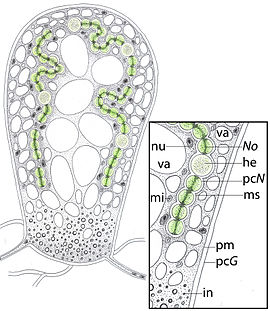
Мицелий гаплоидный , несептированный ( ) или с редкими септами, развивается в почве и проникает в ткани корней растений, где распространяется по межклетникам паренхимы коры . Гифы узкие или широкие (2—10 мкм, иногда до 20 мкм), часто узловатые. У некоторых представителей гифы могут образовывать анастомозы , в том числе анастомозы типа конец к концу, которые соединяют разрушенные соединения гиф. Септы могут образовываться в стареющих частях мицелия или после образования спор .
В отличие от эктомикориз , формирующих наружный чехол вокруг корней, микоризы гломеромицетов практически не изменяют морфологию корней и не сопровождаются утратой корневых волосков . Только иногда на поверхности корня обнаруживается сеть гиф , соединяющих внутритканевой мицелий с грибом, находящимся в почве. Гифы гломеромицетов проникают через клеточную стенку клеток корня и вызывают формирование впячивания плазмалеммы , в котором образуют древовидно ветвящиеся образования — арбускулы, которые обеспечивают физиологические контакты растений и грибов на межклеточном уровне, таким образом, они выполняют питающую функцию. Кроме арбускул, под клеточной стенкой растения-симбионта или за её пределами часто встречаются округлые вздутия — везикулы. В некоторых случаях гриб сосредотачивается в специальных клубеньках. Осуществляемое в ходе симбиоза питание растений происходит при переваривании им арбускул гриба, а иногда везикул и гиф. Наиболее важными из веществ, которые растения получают от грибов в арбускулярной микоризе, являются соединения фосфора . Выделить гломеромицеты в чистой культуре без растений-симбионтов до сих пор не удаётся .
К гломеромицетам относится , внутри вздутий гиф которого располагаются внутриклеточные симбионты — нитчатые цианобактерии из рода Nostoc (как правило, ).
Размножение и жизненный цикл
Большинство гломеромицетов утратили способность к половому размножению . Половой процесс в виде зигогамии описан только у двух видов. В то же время, у Glomus имеется набор из 51 гена , кодирующий белки , необходимые для прохождения мейоза , и предполагается, что виды Glomus могут иметь . Возможно размножение фрагментами гиф, остающимися в почве .
Большинство представителей размножается простыми спорами, формирующимися на концах гиф, диаметром 80-500 мкм () , и более сложными , образующимися в концевых спорангиях . Азигоспоры имеют сложную шестислойную стенку, содержащую хитин и целлюлозу , и служат не только для воспроизведения, но и для перенесения неблагоприятных условий. Споры гломеромицетов всегда многоядерные, они могут содержать от менее 50 до нескольких тысяч ядер . Неизвестно, идентичны ли эти ядра генетически или же они представляют собой смешанный набор генотипов . Они также содержат липидные и белковые глобулы. Они образуются вне корней, реже — внутри них . Споры могут образовываться одиночно, рыхлыми скоплениями, плотными массами или в спорокарпах . Спорокарпы представляют собой агломерации нескольких сотен тысяч спор, а их размер варьирует от менее чем 500 мкм до более 4 см. Спорокарпы иногда покрыты внешним перидием и чаще всего образуются на поверхности земли. Споры могут быть погружены в мицелий или располагаться радиально в переплетении гиф .
Пути прорастания спор различаются в разных таксонах: ростовая трубка может проходить через стенку споры или через место прикрепления к гифе, в процессе прорастания могут участвовать специальные плёнчатые структуры. Прорастание спор может усиливаться под влиянием факторов, продуцируемых растениями. Показано, что стриголактоны индуцируют прорастание спор, оказавшихся вблизи возможного корня-хозяина .
Проникновение гломеромицетов в новые корни может осуществляться посредством спор или непосредственно через гифы, выходящие из колонизированного корня. На поверхности корня гриб формирует аппрессории (гифоподии), которые позволяют гифам проникнуть в клетки ризодермы . Проникновение гиф через покровные клетки и клетки коры направляется и облегчается через формирование растением специального аппарата проникновения .
Гломеромицеты, судя по всему, не отличаются узкой специфичностью в отношении растений-хозяев. Экспериментально показана почти полная совместимость различных видов растений и гломеромицетов. Впрочем, такой вывод был сделан на основании опытов, проведённых в теплицах, а в природе ситуация может оказаться иной. Молекулярные методы анализа также подтверждают отсутствие строгой специфичности. Большинство растений могут образовывать симбиоз с несколькими видами гломеромицетов одновременно, а большинство видов гломеромицетов могут образовывать микоризу с различными видами растениями. Однако некоторые исследования показывают наличие некоторой степени предпочтения хозяина .
Распространение
Связь гломеромицетов с растениями имеет скрытный характер, в связи с чем данных о географическом распространении гломеромицетов практически нет. Некоторые виды были найдены только в одном месте и могут быть эндемиками , в то же время другие распространены повсеместно. Естественным путём распространения гломеромицетов является распространение посредством фрагментов гиф и спорами, переносимыми вместе с частичками почвы. Кроме того, имеются данные о распространении спор гломеромицетов дождевыми червями и млекопитающими . Некоторые спорокарпические виды могут распространяться с помётом грызунов .
Гломеромицеты могут иметь важное хозяйственное значение, так как образование симбиоза с ними может повышать урожайность культурных растений . Например, показано, что гломеромицет при одновременной инокуляции в растение с Trichoderma atroviride действует как биостимулятор, усиливая рост, поглощение питательных веществ и урожайность у овощей .
Классификация
Длительное время грибы, образующие арбускулярную микоризу, рассматривались в составе рода в отделе зигомицетов. Затем было выделено несколько родов, а в 1990 году был описан порядок Glomerales, состоящий из трёх семейств . Согласно филогенетической классификации зигомицетовых грибов 2016 года, грибы, образующие арбускулярную микоризу, относятся к отделу , в котором выделяются в подотдел Glomeromycotina с единственным классом Glomeromycetes . Этот класс включает четыре порядка: Archaeosporales , Glomerales , Diversisporales и Paraglomerales . Самый крупный порядок — Glomerales , а всего в подотделе Glomeromycotina насчитывается около 230 видов .
|
Список выделяемых в настоящее время порядков и семейств гломеромицетов
с указанием числа видов, входящих в их состав |
||
| Порядок | Семейство | Виды |
|---|---|---|
|
Glomerales
|
Glomeraceae
Claroideoglomeraceae |
108
6 |
|
Diversisporales
|
|
10
53 38 7 |
|
Archaeosporales
|
Archaeosporaceae
Ambisporaceae Geosiphonaceae |
2
9 1 |
| Paraglomales | Paraglomeraceae | 3 |
| Семейство incertae sedis | Entrophosporaceae | 3 |
Некоторые исследователи на основании молекулярных и морфологических данных предлагают разделить отдел Glomeromycota на три класса: Glomeromycetes, Archaeosporomycetes и Paraglomeromycetes. В первый класс предлагается включать порядки Glomerales, Diversisporales и новый порядок Gigasporales, в два других — Archaeosporales и Paraglomales соответственно .
Эволюция
Анализ последовательностей генов рРНК малой субъединицы рибосомы (SSU) показал, что гломеромицеты имеют общего предка с высшими грибами (Dikarya) и являются сестринской группой по отношению к аскомицетам и базидиомицетам . Однако данные анализа белоккодирующих генов свидетельствуют о том, что гломеромицеты — одна из групп в составе парафилетической группы зигомицетов . Если гломеромицеты и Dikarya — сестринские таксоны, то объединяющей их чертой является способность вступать в мутуалистические отношения с растениями и водорослями , которая очень редко встречается среди представителей других клад . С зигомицетами гломеромицетов сближает наличие несептированного (ценоцитного) мицелия и ряд черт строения спор и спорокарпиев, которые обе группы могли унаследовать от общего предка .
Исследования с использованием молекулярных маркеров показали, что разнообразие грибов, формирующих арбускулярные микоризы, значительно недооценено, возможно, в связи с тем, что очень многие виды образуют споры очень редко или не образуют их вообще .
Древнейшие ископаемые остатки, достоверно принадлежащие гломеромицетам, представляют собой гломоидные споры и гифы возрастом 460 млн лет, обнаруженные в ордовикском известняке. Таким образом, гломеромицеты появились на Земле раньше сосудистых растений . Возможно, эти древнейшие гломеромицеты вступали в симбиоз с мхами или же с цианобактериями наподобие современного Geosiphon ; возможно, некоторые из них вели сапротрофный образ жизни . Древнейшие и наиболее хорошо сохранившиеся арбускулы имеют возраст 400—412 млн лет; они были обнаружены в корневищах девонских растений, например, . Хотя у этих растений уже имелась проводящая система , однако настоящих корней ещё не было. Таким образом, в процессе эволюции микориза появилась раньше настоящих корней . Стоит отметить, что древние растения вступали в симбиоз не только с гломеромицетами, но также с грибами группы , причём одновременное образование симбиоза с грибами из обеих групп встречается и у современных растений .
Примечания
- , p. 252.
- ↑ , с. 149.
- ↑ , с. 172.
- ↑ , p. 253.
- , с. 62.
- , с. 149—150.
- (неопр.) . Дата обращения: 6 марта 2016. 15 декабря 2017 года.
- Redecker D. , Raab P. (англ.) // Mycologia. — 2006. — Vol. 98, no. 6 . — P. 885—895. — . [ ]
- Gehrig H. , Schüssler A. , Kluge M. (англ.) // Journal of molecular evolution. — 1996. — Vol. 43, no. 1 . — P. 71—81. — . [ ]
- Manfred Kluge, Dieter Mollenhauer, Resi Mollenhauer. // Progress in Botany. — Vol. 55. — P. 130—141. — ISBN 978-3-642-78568-9 . — ISSN . — doi : . 9 июня 2018 года.
- Halary S. , Malik S. B. , Lildhar L. , Slamovits C. H. , Hijri M. , Corradi N. (англ.) // Genome biology and evolution. — 2011. — Vol. 3. — P. 950—958. — doi : . — . [ ]
- Halary S. , Daubois L. , Terrat Y. , Ellenberger S. , Wöstemeyer J. , Hijri M. (англ.) // Public Library of Science ONE. — 2013. — Vol. 8, no. 11 . — P. e80729. — doi : . — . [ ]
- Sanders I. R. (англ.) // Current biology : CB. — 2011. — Vol. 21, no. 21 . — P. 896—897. — doi : . — . [ ]
- Luc Simon, Jean Bousquet, Roger C. Lévesque, Maurice Lalonde. // Nature . — Vol. 363. — P. 67—69. — doi : . 4 марта 2016 года.
- ↑ Arthur Schüβler, Daniel Schwarzotta, Christopher Walker. // Mycological Research. — 2001. — Vol. 105, № 12 . — P. 1413–1421. — doi : . 1 мая 2015 года.
- , p. 253, 258.
- , p. 254.
- , p. 253—254.
- Colla G. , Rouphael Y. , Di Mattia E. , El-Nakhel C. , Cardarelli M. (англ.) // Journal of the science of food and agriculture. — 2015. — Vol. 95, no. 8 . — P. 1706—1715. — doi : . — . [ ]
- Spatafora, J. W. et al. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data // Mycologia . — 2016. — Vol. 108(5). — P. 1028—1046. — doi : .
- Hibbett D. S., Binder M., Bischoff J. F., Blackwell M., Cannon P. F., Eriksson O. E., Huhndorf S., James T., Kirk P. M., Lücking R., Lumbsch H. T., Lutzoni F., Matheny P. B., McLaughlin D. J., Powell M. J., Redhead S., Schoch C. L., Spatafora J. W., Stalpers J. A., Vilgalys R., Aime M. C., Aptroot A., Bauer R., Begerow D., Benny G. L., Castlebury L. A., Crous P. W., Dai Yu-Cheng, Gams W., Geiser D. M., Griffith G. W., Gueidan C., Hawksworth D. L., Hestmark G., Hosaka K., Humber R. A., Hyde K. D., Ironside J. E., Kõljalg U., Kurtzman C. P., Larsson K.-H., Lichtwardt R., Longcore J., Miadlikowska J., Miller A., Moncalvo J.-M., Mozley-Standridge S., Oberwinkler F., Parmasto E., Reeb V., Rogers J. D., Roux C., Ryvarden L., Sampaio J. P., Schüssler A., Sugiyama J., Thorn R. G., Tibell L., Untereiner W. A., Walker C., Wang Zheng, Weir A., Weiss M., White M. M., Winka K., Yao Yi-Jian, Zhang Ning. // Mycological Research. — 2007. — Vol. 111 (Pt. 5). — P. 509—547. — doi : . — . [ ]
- ↑ , p. 251.
- , p. 259.
- Oehl, Fritz; Alves a Silva, Gladstone; Goto, Bruno Tomio; Costa Maia, Leonor; Sieverding, Ewald. // Mycotaxon. — 2011. — Vol. 116, № 15 . — P. 365—379. — doi : . 12 марта 2016 года.
- Arthur Schüβler, Hans Gehrig, Daniel Schwarzott, Chris Walker. // Mycological Research. — 2001. — Vol. 105, № 1 . — P. 5—15. — doi : .
- Redecker D. , Kodner R. , Graham L. E. (англ.) // Science (New York, N.Y.). — 2000. — Vol. 289, no. 5486 . — P. 1920—1921. — . [ ]
- , p. 264.
- Field K. J. , Rimington W. R. , Bidartondo M. I. , Allinson K. E. , Beerling D. J. , Cameron D. D. , Duckett J. G. , Leake J. R. , Pressel S. (англ.) // The ISME journal. — 2015. — doi : . — . [ ]
Литература
- Белякова Г. А., Дьяков Ю. Т., Тарасов К. Л. . Ботаника: в 4 тт. Т. 1. Водоросли и грибы. — М. : Издат. центр «Академия», 2006. — 320 с. — ISBN 5-7695-2731-5 .
- Гарибова Л. В., Лекомцева С. Н. Основы микологии: Морфология и систематика грибов и грибоподобных организмов. — М. : Товарищество научных изданий КМК, 2005. — ISBN 5-87317-265-X .
- Мухин В. А., Третьякова А. С. . Биологическое разнообразие: водоросли и грибы. — Ростов н/Д. : Феникс, 2013. — 269 с. — ISBN 978-5-222-20177-0 .
- The Mycota / Edited by Karl Esser. — Springer, 2014. — Vol. VIIA: Systematics and Evolution. — ISBN 978-3-642-55317-2 .
- 2020-06-16
- 2