
Гора динозавров
- 1 year ago
- 0
- 0


Физиология динозавров исторически была популярным объектом обсуждения у многих палеонтологов и стала причиной множества научных споров, особенно касательно терморегуляции и скорости метаболизма динозавров. В последнее время было найдено много новых доказательств в отношении высокого уровня организации динозавров, опровергающих традиционные представления о физиологии этих животных. Новые данные включают в себя не только свидетельства об особенностях метаболизма и терморегуляции динозавров, но и реконструкцию их дыхательной и сердечно-сосудистой систем.
В ранние годы палеонтологии динозавры представлялись в качестве вялых, громоздких и неуклюжих холоднокровных рептилий . Однако, с открытием более полных скелетов динозавров в западной части США , начиная с 1870-х годов, учёные получили возможность предложить более качественные интерпретации касательно анатомии и физиологии динозавров. Эдвард Дринкер Коуп предложил, что, по крайней мере некоторые динозавры были активными и подвижными животными, как это изображено на картине двух сражающихся дриптозавров , созданной Чарльзом Р. Найтом . Параллельно развитию дарвиновского учения об эволюции , открытие археоптерикса и компсогната привело Томаса Генри Гексли к выводу, что динозавры были тесно связаны с птицами . Несмотря на эти соображения, образ динозавров как больших ящерицеподобных существ уже укоренился в культуре, и большинство аспектов их палеобиологии по-прежнему интерпретировались как типично «рептильные» до конца первой половины двадцатого века . Начиная с 1960-х годов, с появлением фундаментального труда Роберта Т. Беккера « », представления о динозаврах и их физиологии резко изменились. Этому способствовало в том числе и открытие оперённых динозавров в раннемеловых отложениях Китая , поддерживающее связь птиц с мелкими манирапторами .

Изучение динозавров началось в 1820-х годах в Англии . Первопроходцы в этой области, такие, как Уильям Баклэнд , Гидеон Мантелл и Ричард Оуэн , описывали первые и очень фрагментарные находки, принадлежавшие крупным четвероногим животным. Их ранние работы сегодня демонстрируются в музее , построенном в 1850-х годах, где известные на тот момент динозавры изображаются как своеобразные смеси ящериц и слонов . Несмотря на то, что динозавры были классифицированы как рептилии, Оуэн предположил, что сердца динозавров и их дыхательные системы были похожи на таковые у млекопитающих , а не у современных рептилий .
В конце 1960-х годов вновь возникли идеи о том, что динозавры были организованы не так, как современные рептилии. Все началось с работы Джона Острома , посвящённой дейнониху и эволюции птиц . Его ученик, Роберт Беккер, популяризировал возникшую идею в ряде работ, начиная с « » 1968 года . В своих публикациях Беккер аргументировал идею о том, что динозавры и многие другие, даже более ранние архозавры , были активными теплокровными животными, способными на длительные периоды активности. Беккер также указывал на то, что такого рода представления были широко распространены в конце XIX — начале XX веков, но затем (как и предположения о связи динозавров с птицами) были оставлены — что дало ему основание назвать XX век эпохой «депрессии динозавров», а произошедшую с его участием научную революцию — «Ренессансом динозавров». В апреле 1975 года в журнале Scientific American публикуется статья с соответствующим названием — « ».
В своих работах Беккер использовал множество анатомических и статистических аргументов для доказательства своей точки зрения . Эта методология была отчаянно раскритикована учеными того времени , но начала подтверждаться в современных исследованиях. Эти дебаты вызвали интерес к новым методам определения особенностей палеобиологии вымерших животных, таких как гистология костей, которая была исследована для определения темпов роста многих вымерших архозавров.
Сегодня считается, что многие или, возможно, все динозавры имели куда более высокий уровень обмена веществ, чем холоднокровные рептилии, но также и то, что картина может быть более сложной и разнообразной, чем первоначально предлагал Беккер. В частности, было выдвинуто предположение, что крупные динозавры поддерживали относительно постоянную температуру тела за счёт , или имели промежуточный уровень обмена веществ (так называемую) .

Самые ранние динозавры почти наверняка были хищниками и разделяли несколько общих анатомических черт с ближайшими родственниками, такими как Lagosuchus , в том числе: относительно крупные лезвиеобразные зубы, расположенные в крупных, широко раскрывающихся челюстях и образующие ножницеподобный прикус ; относительно небольшие брюшные полости, поскольку плотоядные животные не нуждаются в крупных пищеварительных системах. Позже хищные динозавры достигали намного более крупных размеров , но сохраняли тот же самый набор признаков. Вместо того, чтобы пережёвывать пищу, эти хищники отрывали куски и проглатывали их целиком, подобно современным крокодилам или хищным птицам .
Пищевые привычки орнитомимозавров и овирапторозавров неочевидны: хотя они развились из хищных теропод , их челюсти сравнительно невелики и у них нет лезвийных зубов типичных хищных динозавров. Также нет никаких доказательств их рациона питания в виде окаменевшего содержимого желудков .
Особенности других групп динозавров четко указывают на то, что они были приспособлены для поедания растительной пищи. Эти черты включают в себя:
Растительноядные завроподы не измельчали пищу, поскольку их зубы и челюсти подходили только для снятия листьев с растений. Птицетазовые динозавры, также травоядные, демонстрируют множество различных адаптаций для потребления растительной пищи. Бронированные анкилозавры и стегозавры имели небольшие головы, слабые челюсти и зубы, и скорее всего кормились подобно завроподам. У пахицефалозаврид были небольшие головы, слабые челюсти и зубы, но отсутствие крупных брюшных полостей предполагает другую диету, состоящую, возможно, из фруктов, семян или молодых побегов, которые были куда более питательными, чем листья . С другой стороны, некоторые орнитоподы , такие, как гипсилофодоны , игуанодонты и различные гадрозавры , имели роговые клювы для отрывания растительности, челюсти и зубы, приспособленные для пережёвывания пищи. Рогатые цератопсы имели сходные адаптации для кормления .
Часто высказывалось предположение, что, по крайней мере, некоторые динозавры заглатывали камни, известные как гастролиты , чтобы измельчать пищу в желудках, и что эту особенность они разделяли с птицами. В 2007 году Оливер Вингс рассмотрел описания гастролитов в научной литературе и обнаружил значительную путаницу, включавшую в себя даже отсутствия согласованного и объективного определения гастролита. Он обнаружил, что проглоченные твёрдые камни или песок могут помочь пищеварению у птиц, которые питаются в основном зерном, но могут быть несущественными для птиц, которые едят насекомых летом и семена зимой, и обычно избавляются от камней и песка летом. Гастролиты часто описывали как важные элементы для зауропод, чей рацион из растительности требовал бы очень тщательного перетирания в желудке. Но Вингс пришёл к выводу, что эта идея неверна: гастролиты встречались лишь с небольшим процентом окаменелостей зауропод; количество найденных гастролитов было небольшим, и во многих случаях камни были слишком мягкими, чтобы эффективно измельчать пищу; большинство гастролитов сильно стёрты и относительно гладкие, но гастролиты, используемые современными животными для измельчения пищи, напротив, как правило шероховаты от износа и разрушены кислотной средой желудка. Поэтому гастролиты зауроподов, вероятно, были проглочены ими случайно. С другой стороны, Вингс пришел к выводу, что гастролиты, найденные вместе с окаменелостями теропод, таких, как Sinornithomimus и каудиптерикс , напоминают таковые у птиц, и что использование гастролитов для измельчения пищи могло появиться на ранней стадии развития динозавров, ведущей к птицам .
При откладывании яиц у самок образовывался особый тип костной ткани в конечностях между твердой наружной частью кости и костным мозгом . Эта так называемая , накапливающая кальций, который используется для создания яичной скорлупы. Эта кость рассасывается у современных птиц после того, как они отложили яйца . Медуллярная кость была обнаружена в окаменелостях теропод тираннозавра и аллозавра , а также у орнитопода Tenontosaurus .
Поскольку эволюционная линия динозавров, которая включает аллозавра и тираннозавра, отделилась от линии, ведущей к Tenontosaurus ещё на заре эволюции динозавров, наличие медуллярной кости в обеих группах свидетельствует о том, что, скорее всего, все или большинство динозавров вырабатывали медуллярную костную ткань. С другой стороны, крокодилы, которые являются вторыми ближайшими родственниками динозавров после птиц, не производят медуллярную костную ткань перед откладыванием яиц. Эта ткань, возможно, впервые появилась у базальных представителей орнитодир (группы архозавров, включающей две эволюционные линии: динозавроморфов и птерозавроморфов ), предшествующих динозаврам .
Медуллярная кость была обнаружена у подростковых особей, что говорит о том, что динозавры достигали половой зрелости до того, как они вырастали полностью. Половая зрелость до достижения полного размера встречается у современных рептилий, а также у млекопитающих среднего и крупного размера, однако птицы и мелкие млекопитающие достигают половой зрелости только после того, как они полностью выросли. Ранняя половая зрелость обычно связана с особенностями жизненного цикла животных: молодые особи рождаются относительно хорошо развитыми и самостоятельными, а не беспомощными; а смертность среди взрослых может быть довольно высока .
С 1870 года ученые в целом согласились с тем, что посткраниальные скелеты многих динозавров имеют большое количество заполненных воздухом полостей, которые особенно распространены в позвонках. Пневматизация черепа (например, придаточных пазух ) встречается как у синапсид , так и у архозавров, но посткраниальная пневматизация встречается только у птиц, не-птичьих динозавров и птерозавров. Долгое время эти структуры рассматривались как простые полости для снижения веса, но Беккер предположил, что они могут быть связаны с воздушными мешками, напоминающими по структуре те, что присутствуют у современных птиц . Среди современных позвоночных птицы обладают уникальной по сложности дыхательной системой, характеризующейся сочетанием небольших слабо растяжимых лёгких, вместительных лёгочных воздушных мешков, которые образуют полости в костях посткраниального скелета (то есть пневматизацию), однонаправленной вентиляцией лёгких и эффективного перекрёстного газообмена. Крокодилы, другие современные архозавры, также обладают однонаправленной вентиляцией лёгких, но не имеют воздушных мешков и пневматизации посткраниального скелета . Подобная однонаправленная вентиляция с перекрёстным газообменом позже была обнаружена и у представителей лепидозавров , ведущий активный образ жизни и несколько отдаленных от типичного «рептильного» уровня организации — варанов .

Джон Рубен — один из основных критиков «Ренессанса динозавров» и приверженец традиционных взглядов, вместе с другими авторами (работы 1997, 1999, 2003 и 2004 года) оспаривали наличие у динозавров воздушных мешков и предположили, что у динозавров была дыхательная система вроде той, что наблюдается у современных крокодилов и приводится в действие своеобразным печеночным поршнем — мышцей, прикрепленной в основном к лобковой кости и перемещающей печень назад, что заставляет легкие расширяться, чтобы вдыхать кислорода; а когда эти мышцы расслабляются, легкие возвращаются к своим прежним размерам и форме, и животное выдыхает. Они также предоставили свои аргументы как причину для сомнения в том, что птицы происходят от динозавров .
Критики утверждают, что довольно скромные модификации дыхательной и кровеносной систем, позволили бы современным рептилиям достичь от 50 % до 70 % от объёма потока кислорода, проходящего через легкие млекопитающих подобного размера , а также что отсутствие воздушных мешков не помешало бы повышению уровня метаболизма у динозавров . Однако, основным аргументом Джона Рубена и др. в пользу наличия у динозавров печеночно-поршневого механизма вентиляции легких, являются окаменелые останки Sinosauropteryx с отпечатками мягких тканей, которые были сильно деформированы. Дальнейшие исследования показали, что по ним невозможно было определить, сохраняет ли печень правильную форму, и способна ли она действовать как часть печено-поршневого механизма . Некоторые авторы до сих пор упоминают точку зрению Рубена и др., но не дают каких-либо своих комментариев .
В дальнейшем различные авторы предоставили убедительные доказательства и аргументы в пользу наличия воздушных мешков у завроподов , прозавроподов , целурозавров , цератозавров и тероподов Aerosteon и Coelophysis . У продвинутых зауроподов (« неозауроподов ») задние спинные позвонки демонстрируют явные признаки наличия воздушных мешков. У ранних зауроподов только шейные позвонки имеют эти особенности . Теропод Coelophysis происходит из позднего триаса и является одним из самых ранних динозавров, чьи окаменелости демонстрируют наличия каналов для воздушных мешков . Aerosteon , позднемеловой мегарапторид, имел наиболее птицеподобные воздушные мешки, известные для любых не-птичьих динозавров . Ранние зауроподоморфы — представители группы, традиционно называемой «прозауроподам», также могли иметь воздушные мешки. Хотя возможные пневматические углубления были обнаружены у Plateosaurus и Thecodontosaurus , они были очень малы. В одном из исследований 2007-го года указывается, что у прозауроподов, вероятно, были брюшные и шейные воздушные мешки, что основано на доказательствах их присутствия у сестринских таксонов (тероподов и зауроподов), хотя неизвестно, какое строение имели их легкие . Ещё один аргумент в пользу присутствия воздушных мешков и их участия в вентиляции легких у прозауроподов связано с реконструкцией объёма воздухообмена (объема воздуха, перемещаемым с каждым актом дыхания) у Plateosaurus , который должен был иметь отношение объёма вдыхаемого воздуха к массе тела около 29 мл/кг, что сходна с величинами у гусей и других птиц, но намного выше, чем у типичных млекопитающих .
Вычисление объёмов различных частей дыхательной системы зауропода Apatosaurus целом подтверждает наличие птичьих воздушных мешков у зауроподов:

Исходя из этого, у апатозавра не могло быть «рептильной» дыхательной системы, так как в таком случае дыхательный объём был бы меньше объёма мёртвого пространства. Аналогично, дыхательная система млекопитающих обеспечивала бы легкие лишь около 41 л свежего, насыщенного кислородом воздуха при каждом вдохе, чего было очень мало. Следовательно, у апатозавра должна была быть дыхательная система, неизвестная для современных животных, либо как у птиц, с несколькими воздушными мешками и однонаправленной вентиляцией легких. Кроме того, при птичьей дыхательной системе с воздушными мешками, апатозавру хватило бы легких с объёмом около 600 литров, в то время как при дыхательной системе как у млекопитающих апатозавру потребовался бы объём легких около 2950 литров, что превысило бы приблизительно 1700 литров пространства, доступного в грудной клетке апатозавра .
До сих пор не было обнаружено никаких явных признаков наличия воздушных мешков у птицетазовых динозавров (данные ограничены неявными свидетельствами возможного наличия неинвазивных воздушных мешков у Scelidosaurus и Mantellisaurus , и одна неубедительная находка предполагаемого пневматического отверстия в ребре игуанодонта Delapparentia ). Однако, предполагается, что у них могли присутствовать воздушные мешки, не образующие полости в костях, либо циркуляция больших объёмов воздуха в легких достигалась иным образом. Воздушные мешки были описаны для птерозавров, что может говорить о том, что некоторые особенности дыхательной системы, присущие современным птицам, могли появиться ещё у общего предка птерозавров и динозавров среди базальных орнитодир . Если последовательность развития воздушных мешков у динозавров соответствует последовательности их развития, обнаруженной у эмбрионов птиц, воздушные мешки появились ещё до образования полостей в скелете, которые появляются у более поздних форм ящеротазовых динозавров, но могли не развиться у птицетазовых . Любопытно, что многие триасовые таксоны архозавров и даже развитые архозавроморфы (например, Erythrosuchus ) обладают сложным набором пластинок и углублений на позвонках, которые скорее всего гомологичны аналогичным структурам у орнитодир. Они не связаны с полостями в костях и, как показано в более ранних работах, не могут считаться настоящей пневматизацией скелета и однозначно доказывать развитие у этих животных системы воздушных мешков. С другой стороны, система небольших неинвазивных лёгочных воздушных мешков, которая теоретически могла присутствовать у этих таксонов, может быть базальным признаком для архозавров в целом, и быть вторично утраченной у круротарзов или, по крайней мере, у крокодилов .
В целом было предложено три объяснения для развития воздушных мешков у динозавров :
Дыхательные системы динозавров с птичьими воздушными мешками, возможно, были способны поддерживать более высокий уровень активности, чем тот что доступен для млекопитающих аналогичного размера. В дополнение к очень эффективному снабжению крови кислородом, быстро циркулирующий воздушный поток был бы эффективным механизмом для охлаждения, который необходим для животных, которые ведут активный образ жизни но имеют слишком крупные размеры, чтобы избавляться от избыточного тепла через их шкуры .
Палеонтолог Питер Уорд считает, что эволюция системы воздушных мешков, которая имелась даже у самых ранних динозавров, возможно, изначально была ответом на очень низкое (11 %) содержание кислорода в атмосфере в триасовом периоде .
У птиц имеются костяные выросты, исходящие от вертикального сегмента каждого ребра в каудальном направлении, и придающие грудным мышцам большее количество рычагов для улучшения подачи кислорода. Размер крючковидных отростков ребер связан с образом жизни птиц и вдыхаемым количеством кислорода: они наиболее короткие у ходящих птиц и самые длинные у плавающих, которые должны быстро пополнять запасы кислорода при всплытии. У не-птичьих тероподов манирапторов также были обнаружены крючковидные отростки ребер, по пропорциям напоминающие те, что присутствуют у современных ныряющих птиц и указывают на высокие кислородные требования .
Отростки, которые, возможно, функционировали аналогичным образом, наблюдались у орнитишии Thescelosaurus и были интерпретированы как свидетельство высокого потребления кислорода и, следовательно, высокой скорости метаболизма .
Носовые (или респираторные) турбиналии представляют собой свёрнутые структуры из тонкой кости в полости носа. Они присутствуют у большинства млекопитающих и птиц и выстланы слизистыми оболочками, которые выполняют две функции: они улучшают обоняние путём увеличения площади для анализирования запахов, а также нагревают и увлажняют вдыхаемый воздух, забирают тепло и влагу из выдыхаемого воздуха, чтобы предотвратить высыхание лёгких.
Джон Рубен и другие критики утверждали, что у динозавров и других архозавров (например, наземных крокодиломорфов), для которых предполагается теплокровность, не было обнаружено никаких признаков наличия носовых турбиналий. У всех динозавров, которых они исследовали, носовые проходы были слишком узкими и короткими, чтобы вмещать носовые турбиналии, и поэтому с их точки зрения динозавры не могли бы выдерживать скорость дыхания, необходимую для обеспечения уровня обмена веществ как у млекопитающих или птиц во время отдыха, иначе их лёгкие были бы высушены . Однако, против этого аргумента были выдвинуты серьёзные возражения. Назальные турбинали отсутствуют или очень малы у некоторых птиц (например, Palaeognathae , Procellariiformes и Falconiformes ) и млекопитающих (например, китообразных , муравьедов , летучих мышей , слонов и большинства приматов ), хотя эти животные безусловно теплокровны и в некоторых случаях очень активны . Дыхательные системы некоторых эктотермных животных, таких как ящерицы (в особенности — вараны и тегу , имеющие очень высокие аэробные способности ), также пропускают большое количество кислорода, но отсутствие носовых турбиналий не приводит к высушиванию лёгких . Другие исследования заключают, что носовые турбиналии хрупки и редко встречаются в виде окаменелостей. В частности, они не обнаружены у ископаемых птиц .
В 2014 году Джейсон Бурк и соавторы в журнале Anatomical Record сообщили об обнаружении носовых турбиналий у пахицефалозавров . Кроме того, это кости были описаны ещё в 1977 году у анкилозаврида .

Вполне ожидаемо, что у динозавров будут четырёхкамерные сердца, способные обеспечить достаточное кровяное давление для того, чтобы транспортировать кровь по вертикали на расстояние более одного метра от сердца . По всей видимости, четырёхкамерное сердце является базальным признаком кровеносной системы у всех архозавров, поскольку оно присутствует как у крокодилов (у которых оно, тем не менее, вторично изменено для понижения уровня обмена веществ ), так и у птиц . В настоящее время считается, что четырёхкамерное сердце синапсидов и архозавров развилось независимо от допускающего смешивания артериальной и венозной крови трехкамерного сердца лепидозавров, черепах и земноводных (которые, в свою очередь, также развились независимо друг от друга), и все эти структуры берут свое начало от сердец кистеперых рыб, скорее всего, имевших четыре не полностью разделенные камеры (аналогичная структура присутствует у примитивных неотенических земноводных из семействе сиреновых ) . Любопытно, что у варанов, питонов и наземных черепах (у некоторых из которых в свою очередь известны случаи проявления факультативной эндотермии или мезотермии) сердце функционально приближено к четырёхкамерному, почти полностью исключая смешивание артериальной и венозной крови . Однако, поскольку у предков этих животных некогда развилось трехкамерное сердце, разделение артериальной и венозной крови у них происходит не так эффективно, как у животных с изначально четырёхкамерными сердцами, у которых оно полностью отсутствует без наличия дополнительных модификаций (как в сердце у крокодилов) .
В 2000 году был описан скелет динозавра Thescelosaurus , который имеет то, что было идентифицировано как окаменелые остатки четырёхкамерного сердца и аорты. Авторы интерпретировали эту структуру сердца как указание на повышенную скорость метаболизма и теплокровность у тесцелозавра . Однако, их идентификация была оспорена; другие исследователи опубликовали статью, в которой утверждали, что «сердце» в действительности является простым куском минерала. Они отмечают, что анатомия этого объекта неверна, и аналогичное минеральное образование находится за правой ногой . Однако, авторы оригинального описания отстаивали свою позицию; они согласились, что образование содержит часть минеральных включений, но это не относится к структуре, которая по их словам частично сохранила некоторые части сердца и аорты .
Независимо от идентификации этого объекта, это может иметь мало общего со скоростью метаболизма, поскольку четырёхкамерное сердце лишь позволяет животному развить высокий уровень обмена веществ, но не обязательно означает его наличие .
Яйца динозавров относительно невелики и даже развитые эмбрионы крупных динозавров были найдены в относительно маленьких яйцах (например, Maiasaura ) . Динозавры любых размеров росли относительно быстрее, чем современные холоднокровные рептилии; но ближайший аналог от теплокровных животных по скорости роста зависит от конкретных размерных категорий :
| Вес (кг) | Скорость роста | Современные животные из этой размерной категории |
|---|---|---|
| 0,22 | Медленнее сумчатых . | Крыса |
| 1 — 20 | Подобно сумчатым, медленнее чем выводковые птицы . | От морской свинки до андского кондора |
| 100- 1000 | Быстрее сумчатых, подобно выводковым птицам, медленнее чем у плацентарных млекопитающих . | От красного кенгуру до полярного медведя |
| 1500 — 3500 | Подобно большинству плацентарных млекопитающих. | От гаура до белого носорога |
| 25000 и более | Подобно китам, но медленнее, чем у гипотетической птенцовой птицы , увеличенной до таких же размеров. | Крупные киты |
Согласно исследованию 2004 года , Tyrannosaurus rex достигал массы около 500 кг в возрасте 10 лет, после чего рос очень быстро до 20 лет (набирая более 500 кг в год), где достигал массы, по самым консервативным оценкам, не менее 5-6 тонн, после чего рост значительно замедлялся . В 2008 году исследование одного скелета гадрозавра Hypacrosaurus показало, что этот динозавр рос ещё быстрее, достигая полного размера в возрасте около 15 лет. Авторы обнаружили, что это согласуется с теорией жизненного цикла, согласно которой виды-жертвы должны расти быстрее, чем их хищники, если они теряют много молодняка в результате хищничества, а местная среда обеспечивает достаточное количество ресурсов для быстрого роста .
Основной проблемой оценки скорости роста динозавров является то, что так называемые « годичные кольца » на срезе костей на самом деле могли закладываться чаще, чем один раз в год, что свойственно современным эндотермам, и таким образом нивелировать все получаемые в подобных исследованиях результаты . Кроме того, большой диапазон оценок массы тела динозавров, приводимый в различных работах, может существенно повлиять на ежегодное увеличение массы растущего животного при оценке скорости роста.
С момента открытия динозавров в середине 19-го века, мнение об их образе жизни, метаболизме и терморегуляции со временем изменилось. Активность метаболических ферментов напрямую зависит от температуры, поэтому контроль температуры тела жизненно важен для любого организма, будь то холоднокровное или теплокровное животное. Животные могут быть классифицированы как пойкилотермные (холоднокровные), которые устойчивы ко внутренним колебаниям температуры, и гомойотермные (теплокровные), которые должны поддерживать постоянную температуру тела. Далее организмы могут быть классифицированы как эндотермы, которые регулируют температуру тела внутреннее, и эктотермы, которые регулируют температуру тела за счет использования внешних источников тепла. «Теплокровность» — сложный термин со множеством значений, поскольку в разном контексте он может включать в себя как некоторые, так и все признаки из следующих (и, соответственно, включать или исключать из определения разные организмы):
Предполагается, что крупные динозавры могли поддерживать относительно постоянную температуру тела за счет инерциальной гомойотермии. Теплоемкость столь крупных животных может быть настолько высокой, что им потребовалось бы два дня или более, чтобы температура их тела значительно снизилась при понижении температуры окружающей среды, и это сглаживало бы колебания температуры тела, вызванные ежедневными температурными изменениями, не требуя выработки тепла за счет эндотермических реакций. Инерциальная гомойотермия в настоящее время наблюдается у крупных крокодилов. Однако, рассчитано, что этот механизм был бы неэффективным для динозавров, имеющих массу менее 700 кг. Поддержание постоянной температуры тела за счет инерциальной гомойотермии было бы невозможным для мелких видов динозавров и молоди крупных видов . Ферментативное расщепление растительности в кишечниках крупных травоядных также может привести к выделению большого количества тепла, но этот метод поддержания высокой и стабильной температуры не был бы возможен для хищных динозавров, мелких травоядных или молоди более крупных травоядных . Таким образом инерциальная гомойотермия едва ли была важным методом сохранения тепла, по крайней мере у мелких динозавров (у крупных, с другой стороны, она могла способствовать сохранению тепла, примерно также, как у современных слонов ).
Поскольку внутренние процессы вымерших животных неизвестны, большинство обсуждений сосредоточено на наличии у динозавров гомойотермии и тахиметаболизма. Оценка скорости метаболизма в свою очередь осложняется различием между показателями во время отдыха и активности. У всех современных рептилий и большинства млекопитающих и птиц максимальные показатели скорости метаболизма во время активности в 10-20 раз превышают минимальные показатели во время отдыха. Однако, у нескольких млекопитающих эти показатели различаются и в 70 раз. Теоретически, наземное позвоночное может иметь «рептильные» показатели скорости метаболизма в состоянии покоя и «птичьи» во время активности. Однако животное с таким низким уровнем обмена веществ при отдыхе не сможет быстро расти. Огромные травоядные зауроподы, возможно, постоянно двигались в поисках пищи, и поэтому их энергетические затраты были бы примерно одинаковыми независимо от того, были ли их показатели метаболизма во время покоя высокими или низкими .
Основные предположения об обмене веществ динозавров таковы :

Роберт Беккер в своем фундаментальном труде пришел к выводу, что динозавры были теплокровными и предполагал, что они унаследовали этот признак от более ранних архозавров. Однако, поскольку эта группа завропсидов существовала более 150 миллионов лет (если не учитывать птиц, которые большинством кладистов рассматриваются как одна из рано отделившихся ветвей манирапторовых теропод), вполне возможно, что некоторые группы развили физиологические особенности, отличающиеся от первоначального состояния и включающие в себя разные уровни метаболизма и особенности терморегуляции. Роберт Рейд в 1997 году предположил, что среди динозавров мог быть распространен промежуточный обмен веществ, или мезотермия , условно назвав их «неудавшимися эндотермами». Он интерпретировал эволюционное развитие эндотермии у архозавров и синапсид вот так: примитивные млекопитающие или развитые терапсиды были вынуждены становиться меньше, поскольку архозавры начали занимать доминирующие ниши в крупных и средних размерных категориях. Уменьшение размеров сделало млекопитающих более уязвимыми для потери тепла, поскольку уменьшило их теплоемкость. Таким образом, это заставило млекопитающих увеличить внутреннее тепловыделение на постоянной основе и, таким образом, стать полноценными эндотермами. С другой стороны, динозавры смогли сохранить промежуточный тип метаболизма, который, на первый взгляд, обеспечивал бы им множество преимуществ: низкий уровень обмена веществ и выработки эндотермического тепла в состоянии покоя, что уменьшило бы их энергозатраты, и позволило бы использовать большее количество питательных веществ для роста; в значительной степени использование инерциальной эндотермии; способность контролировать потери тепла путем расширения и сжатия кровеносных сосудов, как это делают многие современные рептилии; поддержка высокого кровеносного давления за счет четырёхкамерного сердца; высокий уровень метаболизма при активности и вытекающая из него способность в значительной мере полагаться на аэробный метаболизм . Известно, что современные кожистые черепахи , тегу и вараны во время активности способны поднимать температуру тела на более чем 10 °C (у кожистой черепахи, повышающей температуру тела за счет высокой мышечной активности и сохраняющей её за счет слоя подкожного жира — до 18 °C) выше температуры окружающей среды , а у самок питонов выделение большого количества метаболического тепла служит для поддержания температуры инкубации кладки яиц . К другим мезотермным животным, филогенетически более отдаленным от динозавров, относятся сельдевые акулы (хотя обычно у акул значительный подогрев получает только желудок ), некоторые костистые рыбы , такие как тунцы , и отчасти однопроходные млекопитающие . Однако лишь небольшое количество животных в наше время имеет промежуточный тип метаболизма, несмотря на существующий потенциал. Таким образом, вероятно, эта стратегия не является настолько эффективной, насколько может показаться на первый взгляд, и кажется маловероятным, что динозавры могли бы доминировать в крупных размерных категориях 150 млн лет, делая ставку преимущественно на мезотермию .
Арманд де Рикклес, изучив строение костной ткани динозавров обнаружил в ней гаверсовы каналы и решил, что они являются свидетельством эндотермии у динозавров. Эти каналы распространены у теплокровных животных и связаны с быстрым ростом и активным образом жизни, потому что они помогают организму быстро наращивать костную массу в процессе роста и залечивать повреждения, вызванные стрессом или травмами . Плотная вторичная встречается у многих современных эндотермов во время ремоделирования кости, а также у динозавров, птерозавров и терапсидов . Вторичные гаверсовы каналы коррелируют с размером и возрастом, количеством механического стресса и оборотом питательных веществ. Наличие вторичных гаверсианских каналов предполагает сопоставимые скорости роста костей у млекопитающих и динозавров . Беккер утверждал, что наличие фиброламеллярной кости (быстро вырабатываемой и имеющей волокнистый вид) в окаменелостях динозавров также свидетельствует об их эндотермии .
Однако, в результате последующих исследований ряд авторов сочли структуру кости ненадежным показателем метаболизма у динозавров, млекопитающих и рептилий. Кости динозавров часто содержат линии замедленного роста (LAG), образованные чередующимися периодами медленного и быстрого роста; на самом деле, многие исследования подсчитывают кольца роста для оценки возраста динозавров . Формирование колец роста у современных рептилий обычно обусловлено сезонными изменениями температуры, и это сезонное влияние иногда рассматривается как признак медленного метаболизма и эктотермии. Однако, кольца роста встречаются у белых медведей и распространены у млекопитающих, которым свойственна зимовка . Исходя из этих и других примеров, связь между линиями замедленного роста и зависимостью от сезонного роста остается нерешенной . Фиброламеллярная кость довольно распространена у молодых крокодилов и иногда встречается у взрослых особей . Гаверсова кость была обнаружена у черепах и крокодилов, но часто отсутствует у мелких птиц, летучих мышей, землероек и грызунов .
Тем не менее, де Риккле продолжал изучать гистологию костей динозавров и других архозавров. В середине 2008 года он стал соавтором статьи, в которой детально исследовались образцы костной ткани у большего количества архозавров, в том числе ранних динозавров и триасовых наземных крокодиломорфов, и он пришел к выводу, что даже самые ранние архозавры были способны к очень быстрому росту, что свидетельствует о довольно высокой скорости обмена веществ. Хотя сделать выводы о самых ранних архозавроморфах на основании особенностей анатомии более поздних форм сложнее, поскольку очень вероятны видоспецифичные вариации в структуре кости и скорости роста, риск ошибок в анализе был сведен к минимуму . В последующих исследованиях также были обозначены четкие различия между гистологией костей, наблюдаемой у эндотермов от таковой у эктотермов, с учетом множества возможных вариаций, позволяющих с большой точностью определять скорость роста и предполагаемый метаболический статус животного .
Архозавры разделились на три основные группы в течение триаса: орнитодиры и оставались приверженцами быстрого роста; круротарзы также в основном сохраняли быстрый рост, хотя в линиях крокодиломорфов , более близких к современным крокодилам, наблюдалась тенденция перехода к более «рептильным» медленным темпам роста; а большинство других триасовых архозавров имели промежуточные темпы роста .
Эндотермы сильно зависят от аэробного метаболизма и имеют высокий уровень потребления кислорода во время активности и отдыха. Кислород, необходимый тканям, переносится кровью и, следовательно, скорость кровотока и кровяное давление у теплокровных эндотермов значительно выше, чем у холоднокровных эктотермов . Минимальное кровяное давление у динозавров можно оценить измерением вертикального расстояния между сердцем и мозгом, потому что эта колонна крови должна иметь давление в нижней части, равное гидростатическому давлению, рассчитанному исходя из плотности крови и силы тяжести. К этому давлению добавляется то, которое требуется для перемещения крови через систему кровообращения. В 1976 году было указано, что из-за значительного расстояния от сердца до головы у многих динозавров минимальное давление крови уже было в диапазоне такового у эндотермов, и что у них обязательно должно было быть четырёхкамерное сердце, отделяющее поток высокого давления от потока с низким давлением .
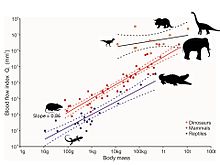
Недавний анализ крошечных отверстий в окаменелых костях конечностей динозавров позволил получить дополнительные данные о скорости кровотока, уже напрямую связанные со скоростью метаболизма. Отверстия называются питательными, а питательная артерия — основным кровеносным сосудом, проходящим через внутреннюю часть кости, где она разветвляется на крошечные сосуды системы гаверсового канала. Эта система отвечает за замену старой костной ткани новой, тем самым восстанавливая микроповреждения, которые естественным образом возникают во время локомоции. Без этого восстановления микроразрушения будут нарастать, что приведет к стрессовым переломам и, в конечном счете, к катастрофическому разрушению кости. В соответствии с уравнением Хагена-Пуазейля размер питательного отверстия обеспечивает показатель проходящего через него кровотока. Разумеется, размер отверстия также связан с размерами тела животного, поэтому данный эффект удаляется анализом аллометрии. Индекс кровотока питательного отверстия бедренной кости у современных млекопитающих увеличивается пропорционально максимальной скорости метаболизма животных, измеренной во время максимальной продолжительной локомоции. Индекс кровотока млекопитающих примерно в 10 раз больше, чем у холоднокровных рептилий. Десять видов ископаемых динозавров из пяти таксономических групп показывают показатели даже выше, чем у млекопитающих, когда учитываются размеры тела, что свидетельствует об очень высокой активности этих животных. Таким образом, высокий уровень кровотока, высокое кровяное давление, четырёхкамерное сердце и устойчивый аэробный метаболизм согласуются с эндотермией динозавров .
Несмотря на то, что они вылуплялись относительно небольшими, динозавры росли очень быстро и могли набрать вес в несколько тонн уже в первые годы жизни. Естественная интерпретация этого факта заключается в том, что молодые динозавры очень быстро переваривали пищу и использовали питательные вещества для роста, что требует довольно быстрого метаболизма и частого кормления . Развивающиеся кости, принадлежащие молодым особям, являются отчетливо пористыми, что связано с васкуляризацией и большой скоростью поступления питательных веществ к кости. Это указывает на темпы роста динозавров, близкие к тем, которые наблюдаются у современных птиц .
Впрочем, некоторые исследования взаимосвязи между размерами взрослых особей, скоростью роста и температурой тела показали, что у крупных динозавров были более высокие температуры тела, чем у меньших; по оценкам, 12979 кг апатозавр, самый крупный динозавр в выборке, имел температуру тела выше 41 °C, тогда как самый маленький динозавр в выборке, 12 кг пситакозавр , по оценкам, имел температуру тела всего лишь около 25 °C. Основываясь на этих расчётах, исследователи сделали вывод о том, что крупные динозавры были инерционными гомойотермами (их температура стабилизировалась за счет размеров) и что динозавры выделяли меньше тепла, чем млекопитающие, по крайней мере пока не перемещались и не переваривали пищу . Температура тела при этом были рассчитана на основе данных об отношении между размерами динозавров и их темпами роста . Однако, темпы роста могут быть связаны не только с уровнем обмена веществ. Исследования зауроподоморфа Massospondylus и раннего теропода Megapnosaurus показывают темпы роста около 3 кг/год и 17 кг/год соответственно, намного медленнее, чем те, которые рассчитаны для майазавра и наблюдаются у современных птиц. Это может быть связано со степенью развитости заботы о потомстве, более высокой у птиц и майязавра .
Однако, сама методология оценки скорости роста динозавров может быть ненадежной, поскольку так называемые « годичные кольца » на срезе костей динозавров на самом деле могут закладываться чаще, чем один раз в год, что свойственно современным эндотермам . Кроме того, оценка скорости роста динозавров напрямую связаны с оценками массы тела, которые могут сильно различаться в работах у разных авторов.
Соотношение изотопов 16 O и 18 O в костях зависит от температуры, в которой находится кость: чем выше температура, тем больше 16 O. В 1999 году было проанализировано отношения изотопов кислорода у двух крупных и высокоразвитых теропод, которые жили в умеренных регионах с сезонные колебания температуры: Tyrannosaurus (США) и Giganotosaurus (Аргентина). Спинные позвонки обоих динозавров не показали признаков сезонной изменчивости, что указывает на то, что температура в глубине тела у них была постоянной, несмотря на сезонные колебания температуры воздуха. Однако, ребра и кости задних конечностей у обоих динозавров показали большую изменчивость в содержании изотопов и более низкую среднюю температуру по мере увеличения расстояния от позвонков. Авторы заключили, что оба динозавра были теплокровными, но имели более низкий уровень метаболизма, чем современные млекопитающие, и что инерциальная гомойотермия была важным фактором терморегуляции у взрослых особей . Аналогичный анализ некоторых позднемеловых птицетазовых динозавров в 1996 году продемонстрировал схожую картину .
Однако, вскоре это мнение было поставлено под сомнение. исследование указывает на гомойотермию, но само по себе не может доказать полноценную эндотермию. Кроме того, развитие костей может быть не непрерывным в области конечностей — у скелетов аллозавридов линии замедленного роста редки или отсутствуют у больших костей конечностей, но встречаются в пальцах. Хотя нет весомых доказательств того, что линии замедленного роста связаны с температурой, они могут отмечать время, когда кости по тем или иным причинам переставали расти. Если это так, то данные об соотношении изотопов кислорода были бы неполными, поскольку этот метод ненадежен для оценки температур, если нельзя показать, что рост костей был одинаково непрерывным во всех частях животного.
Беккер утверждал, что хладнокровные хищники нуждаются в гораздо меньшем количестве пищи, чем теплокровные, и поэтому одинаковая биомасса жертв может поддерживать большее количество холоднокровных хищников, чем теплокровных. При этом отношение общей массы хищников к общей массе добычи в экосистемах динозавров, более ранних архозавров и терапсидов было ближе к соотношению у современных или недавно вымерших экологических сообществ с теплокровными хищниками, чем отношение современных или вымерших экологических сообществ. с холоднокровными хищникам. Это может указывать на то, что хищные динозавры были теплокровными. И поскольку самые ранние динозавры (например, Staurikosaurus и Herrerasaurus ) были хищниками, все динозавры изначально должны были быть теплокровными .
Этот аргумент подвергся критике по нескольким причинам, и не рассматривается некоторыми авторами. Оценки веса динозавров широко варьируются, и даже небольшое изменение могут существенно повлиять на рассчитанное соотношение хищников и их жертв. Беккер получил свои оценки, сославшись на музейные образцы, но они имеют предвзятость к редким или хорошо сохранившимся экземплярам и не отображают полной картины фауны, сохранившейся в палеонтологической летописи. Даже сами ископаемые слои могут не точно передавать численность фактических популяций из-за условий сохранения и неполноты палеонтологической летописи. Для крупных эктотермных хищников, за исключением комодского варана , нет качественных данных о соотношении численности хищников и их добычи. Крупные эктотермные растительноядные в наше время и вовсе отсутствуют. По этой причине Беккеру пришлось сравнивать хищник-жертва соотношение млекопитающих с таковым у рыб, мелких рептилий и беспозвоночных, где ожидаемая продолжительность жизни организмов намного меньше, и другие различия могут также искажать сравнение. Этот аргумент предполагает, что популяции хищников ограничены только наличием добычи, хотя существуют другие факторы, такие как нехватка мест гнездования, каннибализм, конкуренция или хищничество со стороны других хищников, которые могут удерживать популяции хищников ниже предела, налагаемого биомассой жертвы. Неизвестны многие экологические факторы, которые могли повлиять на соотношение численности хищников и их жертв в сообществах динозавров: некоторые хищники могут специализироваться на определённых видах жертвы, в том числе на разных этапах роста; на численность жертв могут влиять болезни, паразиты и голод, которые снизят их численность прежде чем хищники получат возможность охотиться на них .

Конечности динозавров располагались парасагиттально, то есть, находились под прямым углом относительно их тел, также как у птиц и большинства млекопитающих, и не были расставлены в стороны, как у типичных холоднокровных животных вроде ящериц и тритонов . Свидетельством тому являются углы поверхностей суставов, а также точки прикрепления мышц и сухожилий на костях. Все ранние попытки реконструировать динозавров с расставленными в стороны конечностями приводили к реконструкциям с вывихнутыми бедрами, коленями, плечами и локтями .
Существует так называемое ограничение Карриера , которое гласит, что дышащие атмосферным воздухом позвоночные, которые имеют парные легкие и при движении изгибают свои тела в бок, имеют трудности с тем, чтобы передвигаться и дышать одновременно. Это сильно ограничивает выносливость и заставляет их тратить больше времени на отдых, чем на перемещение. Располагающиеся по бокам тела конечности требуют бокового изгибания во время локомоции (за исключением черепах, подвижность тела которых ограничена панцирем, сильно снижающим мобильность). Однако, несмотря на ограничение Карриера, располагающиеся по бокам конечности с энергетической и функциональной точки зрения выгодны для холоднокровных животных, которым не свойственны длительные периоды активности; в частности, они минимизируют время и энергозатраты, необходимое на то, чтобы подняться на ноги из лежачего положения. Парасагиттальные конечности увеличивают затраты и время, необходимое на то чтобы лечь или встать, но являются одним из способов обхода ограничения Карриера и общего повышение скорости передвижения, также требующего больших энергозатрат .
Таким образом, строение конечностей динозавров указывает на то, что они были активными животными, поскольку в противном случае им были бы не выгодны выпрямленные конечности. Активный образ жизни в свою очередь требует высокой скорости метаболизма, который позволяет в значительной степени использовать аэробный метаболизм. Другие доказательства эндотермии динозавров включают в себя длину конечностей (многие динозавры обладают сравнительно длинными конечностями, адаптированными к быстрому бегу) и бипедализм, который в наше время встречаются только у эндотермных животных . Многие бипедальные динозавры обладали грацильными костями конечностей с коротким бедрами относительно длины голени. Это, как правило, является адаптацией к активному бегу, характерному для эндотермов, которые, в отличие от эктотермов, способны продуцировать достаточную энергию в ходе активности, и уменьшить пагубное влияние анаэробного метаболизма на мышцы .
Баккер и Остром указывали на то, что у всех динозавров были прямые задние конечности и что у всех квадропедальных динозавров были прямые передние конечности; и что среди живых животных только теплокровные млекопитающие и птицы имеют прямые конечности (Остром признал, что полупарасагиттальное положение конечностей у крокодилов было частичным исключением). Баккер утверждал, что это было явным доказательством эндотермии у динозавров, в то время как Остром считал его весомым, но не достаточно убедительным .
Исследование, проведенное в 2009 году, подтвердило гипотезу о том, что эндотермия была широко распространена у, по крайней мере, крупных нептичьих динозавров, и что она была бы правдоподобна даже для недалёких предков динозавров, исходя из биомеханики их бега .

В условиях резкой смены температуры, теплокровные животные как правило развивают изолирующие покровы, такие как шерсть, перья или подкожный жир. В настоящее время нет никаких сомнений в том, что у многих видов тероподов были перья, включая Shuvuuia , Sinosauropteryx и Dilong . Они были интерпретированы как изолирующие покровы и, следовательно, свидетельствуют о теплокровности .
Но отпечатки от перьев были найдены только у целурозавров, и поэтому в настоящее время перья, как разновидность изолирующих покровов, не дают никакой информации об эндотермии у других крупных групп динозавров. На самом деле, окаменелые отпечатки кожи карнозавров (таких как Allosaurus и Carnotaurus ), а также очень крупных целурозавров (например, тираннозавридов ) демонстрируют чешуйчатую кожу . Тем не менее, все эти динозавры имеют массу взрослых особей более 1 тонны, и млекопитающие подобных размеров могут иметь короткую шерсть, либо не иметь её вовсе (например, слоны и носороги ). Крупные животные имеют большую теплоемкость и могут сохранять тепло за счет своих крупных размеров. В условиях мягкого климата без резкой смены температур возможно существование и менее крупных эндотермов, не имеющих эффективных изолирующих покровов (к примеру, люди и бабируссы ).
Отпечатки кожи Pelorosaurus и других зауроподов демонстрируют крупные чешуи, а некоторые другие зауроподы, такие как Saltasaurus , имели ещё и костяные пластины. Кожа цератопсов состояла из крупных многоугольных чешуек, иногда с рассеянными круглыми пластинами . «Мумифицированные» остатки и отпечатки кожи от гадрозаврид также демонстрируют чешуйчатые покровы чешуйки. Маловероятно, что анкилозавриды, такие как Euoplocephalus , имели изолирующие наружные покровы, так как большая часть площади их тела была покрыта остеодермами. Точно так же нет никаких доказательств изолирующих покровов у стегозавров .Таким образом, появление изолирующих покровов, возможно, было ограничено некоторыми тероподами (хотя у других динозавров могли имели место быть иные механизмы сохранения тепла, включая крупный размер или даже подкожный жир). Стоит упомянуть, что волокна, похожие на перья, были обнаружены у трех видов мелких птицетазовых динозавров: Psittacosaurus , Tianyulong и Kulindadromeus , но на данный момент они не могут быть классифицированы в качестве перьев.

Окаменелости динозавров были найдены в районах, которые были близки к полюсам в те периоды времени, особенно в юго-восточной Австралии, Антарктиде и северной части Аляски. Нет никаких свидетельств серьезных изменений положения оси Земли, поэтому полярным динозаврам и остальным представителем тех экосистем приходилось справляться с экстремальным изменением продолжительности светового дня в течение года, которые происходят сегодня в аналогичных широтах .
Исследования ископаемой растительности свидетельствуют о том, что в северной части Аляски максимальная среднегодовая температура составляла 13 °C, а минимальная среднегодовая температура варьировалась от 2 °C до 8 °C в последние 35 миллионах лет мелового периода. Ожидаемо, что в этих местах не было найдено окаменелостей крупных хладнокровных животных, таких как ящерицы и крокодилы, которые были распространены южнее. Это говорит о том, что по крайней мере некоторые нептичьи динозавры были адаптированы к холодному климату, для чего необходима теплокровность. Было также предложено, что полярные динозавры, возможно, мигрировали в более теплые регионы по мере приближения зимы, что позволяло им населять столь холодные места обитания даже будучи холоднокровными . Однако, столь длительные миграции требовали бы большего количества энергии, чем аккумулируют холоднокровные наземные позвоночные . В статье 2008 года о миграции динозавров авторы предположили, что большинство полярных динозавров, включая тероподов, зауроподов, анкилозавров и гипсилофодонтов, вероятно, зимовали, хотя гадрозавриды, такие как Edmontosaurus , будучи теплокровными были, вероятно, способны мигрировать на расстояния до 2600 км .
Труднее определить климат в юго-восточной части Австралии, от 115 до 105 миллионов лет назад. Эти отложения свидетельствуют о вечной мерзлоте и бугристой почве, образованной движением подземного льда, что предполагает среднюю годовую температуру от −6 °C до 5 °C; Исследования изотопов кислорода в этих отложениях дают среднюю годовую температуру от 1,5 °C до 2,5 °C. Однако разнообразие ископаемой растительности и большие размеры некоторых ископаемых деревьев превышают то, что наблюдается в таких холодных средах сегодня. Ежегодная миграция от юго-восточной части Австралии и обратно была бы очень сложной для довольно мелких динозавров, таких как Leaellynasaura , потому что на севере переход к более теплым широтам блокировали небольшие морские пути . Образцы костей Leaellynasaura и предоставили доказательства, что эти два динозавра имели разные способы выживания в холодных темных зимах: образец Timimus имел линии замедленного роста, и он мог впадать в спячку; но образец Leaellynasaura не имел признаков линий замедленного роста, и поэтому он мог оставаться активным всю зиму . Исследование, проведенное в 2011 году, посвященное костям гипсилофодонтов и тероподов также показало, что эти динозавры скорее всего оставались активными зимой .
Некоторые динозавры, такие как, например, Spinosaurus и Ouranosaurus , имели на спине «паруса», поддерживаемые выростами позвонков. Такие динозавры могли использовать эти паруса для терморегуляции, в том числе для подогрева путем использования парусов в качестве солнечных батарей, или наоборот, для быстрого охлаждения. Но паруса имело очень небольшое количество известных видов динозавров. К тому же, не совсем понятно, у каких динозавров эти структуры действительно образовывали своеобразные паруса и не были скрыты под кожей. Для спинозавра кажется более обоснованной версия о том, что парус служил ему в качестве гиростабилизатора . Терморегуляционная функция также предполагалась для пластин на спинах стегозавров , впрочем, если это могло быть актуальным для стегозавров с большими пластинами, таких как Stegosaurus , то другие стегозавры, такие как Wuerhosaurus , Tuojiangosaurus и Kentrosaurus , обладали гораздо меньшими пластинами с меньшей площадью поверхности. В последнее время сама идея использования пластин для теплообмена у каких-либо стегозавров была поставлена под сомнение .
Поскольку эндотермия позволяет увеличить размеры мозга, требующего большого количества энергии, некоторые авторы полагают, что увеличение размеров мозга указывает на повышенную активность и, таким образом, эндотермию. Коэффициент отношения массы мозга к массе тела (EQ) у различных динозавров варьируется в зависимости от вида. EQ целурозавров часто бывает близок к таковому у млекопитающих и птиц, тогда как другие тероподы и орнитоподы занимают промежуточное положение между птицами и холоднокровными рептилиями, а другие динозавры напоминают в этом плане холоднокровных рептилий .
Исследование, опубликованное Роджером Сеймуром в 2013 году, дало дополнительный аргумент в пользу того, что динозавры были теплокровными. Изучив гребнистых крокодилов , Сеймур обнаружил, что даже если их большие размеры могут обеспечить стабильные и высокие показатели температуры тела, во время активности эктотермический метаболизм крокодилов выдает низкие аэробные способности и при массе тела около 180 кг генерирует только 14 % от общей энергии, генерируемой мышцами эндотермических млекопитающих такого же размера перед полной усталостью. Сеймур сделал вывод, что динозавры должны были бы быть эндотермными, поскольку они нуждались бы лучших аэробных способностях и более высокой суммарной генерации энергии для того, чтобы доминировать над млекопитающими в качестве активных наземных животных в течение всей мезозойской эры . Однако, дальнейшие исследования показали, что в действительности аэробные способности крокодилов и их суммарная генерация энергии в этом исследовании могли быть значительно недооценены .
В июле 2022 года международная команда ученых опубликовала в журнале Science Advances статью, доказывающую способность триасовых динозавров переживать морозы и обитать в холодных регионах Земли. Исследовав следы динозавров на ледовых скалах Джунгарской впадины в Китае, специалисты установили, что ещё до триасово-юрское вымирания некоторые динозавры смогли приспособиться к холодному климату, поэтому смогли выжить и адаптироваться к жизни в полярных областях .

Судя по всему, даже самые ранние динозавры имели анатомические особенности, которые лежат в основе аргументов о теплокровности динозавров. Предложено два объяснения наличия этих признаков у примитивных представителей этой группы архозавров:
Крокодилы представляют собой своеобразную загадку, если сравнивать их с динозаврами как с активными животными с постоянной температурой тела. Крокодилы представляют параллельную линию эволюции архозавров — круротарзов, которые развивались незадолго до динозавров и являются их ближайшими родственниками после птиц. Но современные крокодилы холоднокровны, хотя они и имеют полупарасагиттальные конечности, а также ряд анатомических особенностей кровеносной и дыхательной системы, обычно связываемых с теплокровностью :
В конце 1980-х годов было высказано предположение, что предки крокодилов были активными и теплокровными хищниками, а также то что их предки — архозавры тоже были теплокровными. О возможно вторичной холоднокровности крокодилов вскользь упоминал ещё Роберт Беккер в своей статье «Ренессанс динозавров». Эти идеи также подтверждаются гистологией костей и многими палеонтологическими находками, демонстрирующими что многие крокодиломорфы были легко сложенными, активно бегающими животным и полностью выпрямленными конечностями. Исследования в области эмбриологии показывают, что эмбрионы крокодилов сначала развивают полноценные четырёхкамерные сердца, а затем образуют модификации, которые заставляют их сердца функционировать как трехкамерные при погружении под воду. Используя принцип, что онтогенез повторяет филогенез, исследователи пришли к выводу, что у предков крокодилов были полностью четырёхкамерные сердца и поэтому они могли быть теплокровными, и что позднее некоторые крокодиломорфы претерпели значительные физиологические и анатомические изменения, вернувшись в водную среду обитания. Поскольку экология крокодилов напрямую связана с ожиданием добычи у кромки воды и длительными погружениями, низкий уровень обмена веществ и традиционно считающиеся «примитивными» признаки холоднокровных животных оказались для них выгоднее теплокровности . Однако, многие другие крокодиломорфы не стали возвращаться в водную среду и продолжили развиваться как активные наземные животные наравне с динозаврами . Исследования концентрации изотопов кислорода и гистологии костей (показавшие очень высокие и непрерывные темпы роста) у развитого архозавроморфа Erythrosuchus africanus , базального по отношению к архозаврам (либо представляющего крокодилоподобную ветвь архозавров), также дали убедительные доказательства в пользу того, что это было теплокровное животное, способное на длительными периоды наземной активности . Причем, эти методы исследования указывают на некоторое повышение уровня обмена веществ от стандартного «рептильного» уровня ещё у протерозухий . Если это мнение верно, то развитие теплокровности у архозавров и синапсид заняло сопоставимое количество времени.

















