Белокровные рыбы
- 1 year ago
- 0
- 0

???? ( ???. Pisces) — ???????? ?????? ?????? ????????????? ??????????? ???????? , ????? ??????????? ?????????? . ???? ??????????????? ???????? ???????? ?? ???? ?????? ?????????????????? ???????? ????????? . ? ????? ?????? ?????????? ???????????? ???????? ??????????????????? ????????????? ?????? . ??????, ????????? ??????????? ??????? «????» ????????? ? ???????????????? ?????? , ?? ?????????? ????????????, ? ??????????? ??????????? ???? ??????????? ??? ?????? .
???? ??????? ??? ? ???????, ??? ? ? ??????? ???????? — ?? ???????? ???????????? ?????? ?? ?????? ??????. ???? ?????? ?????? ???? ? ??????????? ?????? ????????? ??? ???????????? ??????? ????? . ?????? ???? ??? ????????????? ????????? ? ???? ? ??????? ????? ?????? ??????????? ????????.
??????????? ???? ????? ??????? ?? 7,9 ?? ( Paedocypris progenetica ) ?? 20 ? ( ??????? ????? ).
? ???? ???????? 35 768 ????? ??? (?? ????????? ?? 2020 ???), ?????? ????????? ??????????? ????? — ?????? ??? ??????????? ????? 300—500 ????? ??? ????? ?????. ? ?????? ??????? ????? 3000 ????? , ? ??? ????? ? ??????? ????? ??????????? ????? 280 ????? .
???????? ??? ???????? ?????? ???????? — ?????????? .
????? «????» ( ??????????? . ???? ) ?????????? ?? ?????????????? * ryba ??????? ?????????? . ?????????????? ? ?.-?.-?. rû??? , rûpa «????????», ????? « ????? », ??.-?.-?. ruppe , rûpe «?????». ?? ??????? , ????? ???????? ?????????????? ????????? ?????? ????? ???????? * z?v? , ??. ???. žuvìs «????», ????. ????? (??. ?????????? — «????? ? ?????») «????», ???. jukn , ???????, ????? ???????? ? ???????? «?????», ?????? ????????? ?? ??????????? .
????????? ?? ???????? ???, pisces , ??. ?. piscis , ??????????, ??-????????, ?? ??????????????????? * pisk- , ?? ???????? ????????? ? ????. fish , ?? ???? «????» .

?????????, ??? ???????? ??????? ???????????? ????????, ???????????? ???, ???????? ?? ??????? ???????? (????? 450—470 ??? ??? ?????). ??? ?? ????? ? 1999 ???? ? ????????? ????????? ??????? ???? ??????? ???????????? ????????????? ???????? Haikouichthys ?? ?????? ???????????? ????????? ????? 530 ????????? ??? (?????? ??????? ). ????????, ???????? ????? ???? ???????? ???? ??????????? .

???????? ?????? ??? ?? ???????????? ????? ???????, ??????????? ????? ?? ???????? ???. ????? ????????, ???? ????? ?????? ????????, ?????????? ??? ? ????? ???????????? ???????? ? ???????? ????. ???????? ?? ????????? ?????? ????????????? ??? ??? ? ????????, ??? ???????? ??????????? ????????? ?????? ?? ?????? . ????? ???????, ???? ? ???????????? ???????????? ?????? 100 ??? ??? ? ???????? ???????????? ????????????, ? ??????? ?? ?????????? ???????. ???????? ???? ????????? ?? ?????? ?????? ? ??????, ????? 420 ??? ??? ?????, ? ???????? ???????? ? ??????? . ???????????? ???? ??????? ? ??????? ??????? ??? ??????? ? ??????; ?? ?????????, ??? ??? ???????????? ??? ? ??????. Guiyu oneiros — ????? ?????? ????????? ???????????? ???? .

????? ???????? ???? ??????? ???? ????? ??????????? , ? ??????, ???????? ???? ?????? FishBase , ????? 31 ?????? ?????, ????????? ? ?????. ????? ?????????? ????? ?????????? ???????? ?????????? ???????? ????? ?????, ? ????? ??????????????? ??????? ????????? ?????. ???? ??????? ???? ???????????? ????? ????????: ???????? ???? (Chondrichthyes), ???????????? (Sarcopterygii) ? ????????? ???? (Actinopterygii). ??? ????????? ?????? ?????????? ?????? ??????? ??? .
??????????? ???????? ??? ????? ?? ??? ?????????: Holocephali (?????????????) ? Elasmobranchii (????????????????, ???? ?????? ????? ? ????? ). ???? ?????????? 900—1000 ????? ???????? ???.
??????? ??????? ??? ???????????? ????? ? ?????? (? ????????? ??? ????? ???????????). ??? ? ? ???? ?????? ???????????, ???? ??? ??????? ?? ????? ? ????????? (??????? ???? ???? ??????????????? ?????????????, ????????? ?? ????????????? ?????). ????????? ? ??? ???????????????. ?????? ? ?????????? ???????????? ?????????????? ?????? (?????), ??????? ???????? ??????? ??????? ?????????.
? ???????????? ????? ???????? ???? ?????? ?????????? ???? ???? — ?????. ???????? ???? ????? ?????????? ?????, ??????? ??????????? ????? ???? ???????????; ??????????? ? ???? ???????? ?? ???????, ?????????? ?????, ??????????, ? ???????????? ? ???? ? ???? ? ??????. ?????????? ????? ??????? ?? ??????? , ??????? ????????? ?????? ?????, ? ?????? ??????? ??????. ?? ??????????? ??????? ?????? ? ????? ???? ????? ? ???????? ? ?????? ????? ????????. ?????????? ?????????? ????? ?? ??????????????, ?? ??? ????? ???? ?? ?????????? ?????????????. ??????????? ???? ????????? ???????? ??? (????????, ? ????????????? ??????? ) ???? ???????? ???????????????? ??????????? ???????.
????????? ????? ??????? ????????? ?????? ????? ?????.

????????? ????? ??????? ? ???????? ??????????? ?? ????????? ???, ???????? ????????? . ??? ???????????? ????????? ???????????, ??????? ?????? ??????? ??????? ???????? ?? ?????? ???????? — ???????; ????? ????? ????? ????????? ???? ???? ???????? ???????? ????????, ??? ? ?????????????? ???????? Polypteridae ? Lepisosteidae .
??? ?????????? ? ??????????? ?????????? ? ????????????? ??? ?????????? ?????????? ????? , ??????? ??????????? ??????? ?????????? ??????? ??????? (?????? ?????????? ????????), ? ????? ???? — ???????. ?????? ???????????? ??????? ???????? ?????. ? ??????????? ????? ?????????? ? ????????????? ??? ??????? ?????????? ? ?????????? ???????? ?????? ?????????? ????????????: ? ??????????? ????? ???? ????????? ?? ??????????? ????? ??????????? ???? ????????? ??????? ???????.
??????? ??? ????? ??????????? ? ????? ??????? ????????: ?? ?????????? ??????????? ???? ????????? ?????? ?? ????????????? «???????????» ???, ????????, ??????????? ????? «???????????????».
??????-???????????? ??????? ??? — ??????? ??????? ? ?????? ???, ??????? ????????? ?? ???????? ???? ????????? ? ?????????? ????? ? ???????????? ????????. ????? ??????-???????????? ???????, ????????? ???????????? ??????????????, ????????????? ????? ? ??? ?????????? ?????? ?????????????????? ???????.
? ??????? ?? ???????? ???????????, ????? ??????? ??????? ?? ????????? ??????, ????? ??? 40 ??????? ????????? ?????? ??? ????? ????????? ??????????. ??? ????????? ?????????? ???????, ?????????? ?? ? ???????, ???????? ???????? ??????? ? ??? ??????? ???????.
????????? ???????? ???????? ? ????? ?????? ???????????? ????????????? , ??????? ???????? ???????? ???? . ???????????? ??????? ??? ? ???????? ????????? ?? ????????? ?????? ???????? ??? , ? ???????? ?????????? ?????? ??????? ?????????.

??????? ? ??????? ??????? ? ???????? ??? ??????????? ???????????? ?? ??????? ???? ???????? ??? (? ??? ??????????????? ????????? ?????? ???? ??? ??? ? ???? — ??? ?????????? ?????? ?????).
? ????????? ??? ??????? ????? ???????? ?????? ????? ?? ???????????????? ( premaxilla ) ? ??????????????? ?????? ( maxilla , ??????? ???????»), ?? dentale ? articulare (?????? ???????).
????????? ?????????????????? ????? ?????? ????????? ??? ??????? ??????? ? ????????? ??????? ? ??????? ?????????? ??????. ???????? ?????????? (???????) ??????????? ???????? ????, ??????? ?????? ?????? ???? ??? ????????? ?????? ??????? ???????. ?? ??? ???? ???????? ???? , ??????? ????? ???????? ??????????? ?????????, ? ???????? ????????? ??????????? ??? ?????????? ????????? ??????? , ??????? ????? ????? ????? ????.
?? ????? ??????? ?????, ??????? ???????? ???????? ?????? ???????, ??????? ???? ???????? ????? ???????, ??? ??????? ??????????? ??????. ??? ???? ? ??????? ??????? ????????? ??????????? ???? ?? ???? ????????? ??? ???. ???????? ?????? ??? ???? ????????? ????? . ????? ?????????? ???????? ???????? ? ?????????? ???? ? ?????????? ???? ? ???.
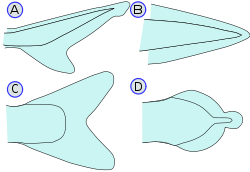
???????? ???? ??? ???????? ??? ????????? ?????????? : ??????? (??????? ? ???????) ? ????????? (???????, ????????, ?????????). ??? ???? ? ??????? ??? ???????? ??????? ?? ??????? (? ????????? ??????????? — ?? ???????? ) ?????, ??????????? ??????????. ?????????????? ? ???????? ????? ????? ????? ????????????? ??? ?????????? ???????, ???????? ??? ?????????? ??? ???????????? ????????????? ????????. ????????? ??????? , ??????? ? ??????????? ??? ???????? ???????? ??????????, ?????????????? ??????? ??????????? ?????????? ?????? ( ???????? ? ??.) ? ??????????????? ? ???? ???? ? ?????????? ? ??????? ?????? ????????. ?? ??????????? ???????? ??????? ? ?????? ??????? ????????? ??????? ????? ???? ?????????????? (????? ??? ??????? ????? ?????? ????????; ??? ?????????? ??? ??????????? ????????? ???) ??? ???????????????? (????? ???? ???????, ?????? ???????, ?????? ??????; ?????????? ??? ???? ? ?????? , ? ????? ????????? ; ? ????? ?????????????? ??????? ??? ??? ????????? ????????? ??????? ???????????????? ? ??????? ?????? ????????).
??????????? ??? ??????? ?? ?????????, ??????????? ????????? . ???????? ??? ??????????? (?? ???? ??? ?? ???????? ??????????? ????????), ????? ?????????? ????????? ???????? ?????????; ?????????? ????, ????????????? ?????? ??? ?????? ?????????, ????????? ??????????? ????? , ?????????? ??????? ???? . ??????????? ??????? ?? ??? ??????: ?????????? ? ?????????. ?? ????????? ???????? ? ??????? ??????? ???????? ????????, ? ??????? ???????? ?????. ? ????????? ?????? ???????????? ??????? ????????? ???, ???? ????? ?????????? ???? ??????? ?????????? (?????????) ????, ??????? ????????????? ? ???????? ????? ? ???????? ?????????? ? ??? ??????? ??????????? ????? — ??????? ????? . ?? ?????????? ? ????????? ??? ??????????? ????? ? ???? ??????? ??????????? ???????? ????????.
???????? ??? ?????? ??????? ??????? ????????? ??????????? ??? ????????? ????????, ? ????? ??? ???????????? ????????? ? ?????????????? ? ????? ???? ? ???? ??? ????? ? ???????? ?? ??????. ? ???????? ???????? ???????? ????????? ? ????????? ??? ?????????? ??? ?????????? ???? ????????? ???????. ???????? ??????? ???????? ??? ???????? ??????? ???? ?? ????????? ???? ??? ????? ????? ???? ????????????? ? ?????? ???????????? ?????? , ???????? ??? ??????????? ??????????????. ?????? ??? ???????? ???????? ? ????????? ????????????? ???-???????? ????? ???? ????????? ? ???????????? ??????? — , ???????? ??? ???????????? ????????????? ?????. ????????? ?????? ? ???????? ????????.
? ???????? ????????? ?????????: ??????? ??????? ( ???. pinna dorsalis , ???????????? ????????? ????????? ?????? D ), ??????? ????? ???? ??????????? ? ?????? ??? ?????? ?? ?????? ?? ????, ?????? ????? ??????????? ?????; , ??? ????????????, ??????? ( ???. pinna analis , ???????????? ?????? A ), ????? ???? ? ?????? ??? ? ?????????? ?? ?????? ?? ????, ??????? ??????????? ?????; ????????? ??????? ( ???. pinna caudalis , ???????????? ?????? C ) ? ????????? ??? ???????????; ??????? ??????? ( ???. pinna adiposa), ????????????? ? ????????? ????? ???.
?????? ????????, ?? ????, ???????????? ????? ?????? ??????????, ??????????? ?? ??????????????? ?? ?????????? ????????? ???????? — ????? ?????????. ? ???????????? ??????????? ??? ???????????? ????? ?????? ?????????: ( ???. pinna pectoralis , ???????????? ????????? ?????? P ) ? ????????? ( ???. pinna ventralis , ???????????? ????????? ????????? ?????? V ). ? ????????? ??? ????? ????????????? ???????, ? ? ?????? ??????? — ? ??????? ????????. ?? ????????? ?? ???????? ?????? ????????? 3 ???????? ???? ??????? ?????????: ????????????? — ??????????? ? ??????? ????? ???? ?????? ??????? ?????????, ??????????? — ??????????? ??? ???????? ??????????, ? ????????? — ??????????? ??????? ??????? ????????? ?? ?????.
? ?????? ? ????? ??????? ?? ???????????? ??????? ???????? ?? ?????????????? ?????, ??????? ?????????? ?????????????? ?????? (????????????) ? ????????? ????? ???? ????, ?????????? ????????? , ?? ?????????? (???????) ? ??????????? (??????) ?????.
???????? ??? ?????????????? ????????? ?????????? ????, ??????? ??????????? ??????????? ? ?????????????. ??????? ? ???? ???? ????? ????? ???????, ????????? ???? ? ??????, ? ??????????? ????????????? ?????????????? ????? (??????????). ?????????? ???????? ????? ????????? ?????????? ?? ???????????, ???????? ??? ? ?????????????? ???????? (?????????) ?? ???? ????? ???? ??? ???? ? ????????? ??? ??????.
? ????? ??????????? ??? ???????????? ????? ?????? ????. «?????????» ????? ???????????? ??? ????????? ????????. ??? ???????? ????? ?????????? , ??????? ????????????? ?? ??????? ????. ?????????? ? ??? ? ???????? ????????, ?? ???? ? ??? ?????????? ?????? ????????? ??????????? ???????. ????? ??????? ????? ????? ????? ?? ?????????? ? ?????? ???????????? ??? ?????? ?????????? ????????. ? ??????? ?? ???????, «???????» ????? ????? ? ??????????????? ?????????????? ???????????? ???????? ? ????????, ?? ???????????????? ??????????. ??? ???????????? ??? ??????? ????????? ??????; ??? ???? ??? ????? ?????? ???????, ??? ??????? ?????, ????????, ?? ?????? ??????????.
? ?????? ??? ????? ????? ????????? ????? ? ????????? ?????? ??????? ????? ????????. ? ????????? ????? ??? ????????? ? ?????????????? (???????????). ? ?????? ( Scombridae ) ????????? ?????????? ??????????? ??????????? ????? ?????????????? ?? ????? ??????? ??????, ??? ??????????? ?????? ?????? ????, ????? ????? ???????? ?? ????????? ? ???????? ???????? ?????.
????????????? ????, ??????? ???????????? ??? ?????????? ????, ???????????? ?????????? ??? ???????????????? ??????; ? ????????????? ?????? ????????????? ???????? , ?????????????? ??????????????? ???????, ???????????? ??? ????????? ?????? ????????. ??????????? ???????? ?????? ??? ?????????? ??????? ????????????? ??????? ??????????? ??????????? ?????????? ? ???????????? ? ?????? ???????? : ??????? ???? ? ???-??????????? ( Uranoscopidae ), ??????????? ??????????? ( ????????????? ????? ) ??? ?????? ??????????? ( ????????????? ???? ).
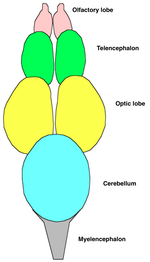
???????? ???? ??? ??????? ?????? ?? ??? ??????? ?????: ????????, ??????? ? ?????? ????. ???????? ???? ??????? ?? ????????? ????? ? ?????????????? ?????. ?? ??????????? (????????) ????? ????????? ????? ??????????? ???????????? ????????, ??????? ???????? ??????? ?? ???????????? ??????????. ???????????? ???????? ?????? ????????? ? ???, ??????? ??????? ?????????? ???, ???????? ? ????. ? ???? ???????? ????? ????????? ?????????? ????. ?????? ????? ?????????????? ?? ?????????? ?????? ???? (? ???? ????????? ???? ? ???????? ) ? ????????????? ???? .
??????? ???? ???????? ?????? ??????? ??? ????????? ?? ???? ????? ???????????? ????. ?????????? ???????? ? ????????????, ? ???????? ???????? ????? ??????????? ???????????. ? ?????? ???????? ???? ????????? ??????? ?????? ? ??????? ???? ????? ?????????? ???????, ? ???????????? ??????? ??????? ?? ???? ????? ???????????.
????? ??? ?? ?????? ???????? ????? ????? ? ??????? ?????? ??????????? . ?????? ??????? ????? ??? ?? ????? ????????????? ??????????? ? ???, ??? ??? ??????????? ???? ?? ???????? ???????? ?????????? , ? ?????????? ??? ? ???????? ??? ???????? ?? ???. ????????? ???????? ? ??? ????????? ? ??????????? ?? ????? ?? ????????: ? ????????????? ????? ????? ????????????? ??? ?????????? ????? ??????????????? ??????? ????? ??????? , ? ????, ??????? ????? ?? ??????????, ???????????? ????? ??????? ??????. ? ????????? ????????????? ???, ???????? ? ???????????????? , ???? ????? ??????????????? ????????, ??????????? ?????????? ? ????????????? ???? (?????????? ????) ?????? ????? ??????????? ????. ????? ????, ? ???? ??? ??????? ?????????? ?????????? «????????? ?????», ?????? ?????????????? ????? ????? ????? ?? ??????????? ?????, ???????, ?????? ????????? ?? ????????? ?????????, ??????? ????????? ?????? ???????? ???? ?????? ???. ??????? ????? ????? ????? ?????? ??? — ??????? ????????????? ??? ???????????? «????????» (?????????) ???, ?????????? ????????? ??? ???????? ??????? ??????. ? 2011 ???? ??? ????????? ????????? ????? ??? ???????? ?????? ??? — Bangana musaei .
???????? ? ???? ????????? ????? ??????????????? ? ?????????? ??????? ?????????? ?????. ??????????? ??? ? ???????? ?????????? ???????? ?????? ???????????? ?????? , ???????, ??? ?? ?????? ?? ???? ? ?????? ????????, ?????????? ?? ?????? ???? ?????? ??? ????? ??? ????, ? ??????? ?????-?? ???? ????? ?? ????. ???????????? ????????? ??? ??????????? ? ???????. ???, ? ??????? ?? ??????? ?????? ???????????, ?? ??????????? ? ??????????? ( ????? ???? ?????? ? ????????????? ???). ???????? ????????? ? ?????? ??? ???? ?? ?????? ? ??????? ???????, ?? ? ?? ???????? ??????????, ?????? ? ???? ????????? ? ?????? ?? ??????????? ????.
??????????????? ??? ?????????? ?? ?????????? ??? (?????? ?????? ????? ? ??????????), ? ????? ??????? ??????? ????? . ?????????? ??? ???????????????? (???? ? ??????) ? ????????? ??? ??????? ?? ???? ??????????? ???????, ????????????? ? ???? ??????? ???????????????? ??????????, ? ???? ?????, ?????? ?? ??????? ???????? ??????? . ????????? ???? ??? (????????, ?????????? ?????? ? ?????? ???? ????? ) ????? ???????? ???????? ( ??????? ??????? ), ??????????? ??? ? ???????????? ??????? . ????????? ???? ????????? ??????? ???????? ??????????? ???????????? ???????, ??? ???????????. ?????? ? ??????? ?????? ???????????? ???? ?????????? ? ????????????.

??????????? ???? ?? ??????????? ???? ????????? ???????????, ??????? ?????????? . ??? ?????? ????? ???? ???????? ?????????? ??? ??????? ??? ?????? ? ???????????? ???????, ??????? ?????????? ??????? ?????? . ?????????? ???????? ????????? ??????? ???????????? (??????) ? ????????? ???????????? ??????, ? ????? ??????? ??????? ???????, ??????? ????????? ?? ???????????? ???????. ??? ???? ???????? ???????????? ??????, ??????? ??????? ????????. ??? ???????? ????????? ????????? ???????? ????????? ??????? ?????????? ?????: ????????? ???? ???, ???????? ????, ??????? ????????????? ? ????????????, ????????? ?????? ?? ?????? ??????? ?????.
???????? ?????????????? ???? — ??????????????? — ??????? ?????? ????? ???, ?? ?????? ???, ??????? ????? ???? ???????????? ????????????? ??????? . ????????????? ??????? ???????????? ? ??????? ??????????? ???? ?? ??????????? ????. ??? ???? ????????? ???????????? ?????????, ??????? ???????? ????????????? ??? ? ???????? ? ???? ?????????????????? ??????, ??????? ???????? ??????? ? ????????? .
?? ????????? ?????????????? ????????????? ????????? ??? ????? ?? ??????????????? (??????????? ???), ?????????????????? ( ??????? ,), ? ??????????????????? ( ????????????? ??? , ????? , ???? ).
??????????????? ???? ? ?????????? ??????-???????? ???????????? ??????? ?????? ????????????? ???????? 100—200 ??. ? ?????? ??? ?????????? ????? ???????????????? ????, ?????????????? ?????????????? ?????????????? ?????? ????????? ??????, ??????? ?????? ?? ????????? ? ?????????? ???. ? ??????????????? ??? (?? ??????????? ???????? , ????????? ????????? , ??????? ) ???????????????? ???????????. ??? ???????????? ????????????? ???, ??????? ????????? ?? ?????? ????????? ? ????????? ??????? ????????? .
?????- ? ??????????????????? ???? ???????????? ????????????? ???? ? ??????? ????????????????? ??????? ?????. ?????????????????? ????????????? ?????? ?????? ?? ??? ??????, ????????? ?? ?????? ? ??????????. ??? ???????????? ????? ??????, ??????????? ????????????? ?? ????? ???? ?????????, ????????? ?? ?????????, ????????? ? ????????. ? ?????? ????????????? ??????, ???????????? ?? 25 % ????? ????, ?????? ?? ???????? ???? . ?????? ????? ??????? ?? ?????????????? 600 ???????????? ?????, ??????? 6 ??????. ?????? ?????? ???????? ???????? 40 ????????????? ??????????? ?????????, ?????????? ??????????? ?????????????? ??????. ?????? ?????? ???????? ????????? ????????????? ???????? .
???????? ???????? ????????????? ??????? ? ??? ???????? ????????????? ???? ?????????????? ????? ? ??????? ???? .
? ??????????? ??? ???? ? ???????????? ??????. ? ????????????? ??? ??? ????? ?????????????? ? ???????? ??????????? ? ?????????? ? ?????????. ??????????? ??????? ?????????, ?????????????? ????? ?? ?????? ????? ????? ? ????? ????. ? ??????? ?? ?????? ?????? ???????????, ?????? ??? ?? ????????????? ??? ????????? (???? ??????????) ??????????? ?????????? ????? (????????????) ?? ????????????? (????????). ?????????? ?????? ??? ???????????? ????? ???????????????? ????? ?? ?????, ??????????? ???????? ??????: ???????? ?????, ?????????? , ????????? ? ???????????? ????? . ?????? ?????? ????????? ?????????, ??????? ????????? ????? ??? ?????????? ?????? ????????? ?????? ? ????? ??????????? (?? ????????? ?????? ? ????????????? ??????), ?? ?? ????????.
???????? ??????? ?????????? ??? ???????? ????? , ??????? ??????????? ?? ???????? ??????? ???????. ? ????????? ??? ??? ??????? ???????? ???????, ? ?????? ??????? — ???????? ?????????????? ??????. ?? ????? ?????????? ???? ???? ???????? ? ??????? ??????? ????? ???, ? ????? ???????? ????? ????????? ?????? ? ??????? ?????? ??-??? ???????? ??????. ???????????? ????? ??????? ?? ??????????????? ??????? ? ??????????? ???????, ??????? ??????????? ?? ??????? ???????? ?????. ????????????? ??????????, ??????????????? ??? ?????????? , ???????? ???????? ????????, ??? ??? ?????? ????????? ????????? ?????? ????????????? ????????? .
? ?????????? ? ?????? ???? ????? ???????????? ? ?????? ??????? ??????????. ?? ?????? ??????? ???????????? ????? ?????????? ?????????????? ????? ????; ????????? ????? ??? ????? «??????», ??? ??????????? ??????????? ?????? ( ???? ); ????????? ???? ????? ?????? ???????? ??????????????? ( ?????? ).
???? ???? ??????????? ? ?????????? ?????? (?????? ?????????????????????), ???????????? ?? ???. ??? ??? ????? ?????? ? ????? ????? ??????? ???? ???????? ? ???????, ??? ???????????? ????????? ??????????, ????????????? ? ?????????? ????. ?????? ????? ?????????? ??????? ? ?????? ????? ? ?????? ??? ??????? ????? ?????????????? ??????? — ???????????? ???????? , ??????? ?????? ??? ?????????? ??????????????? ???????????, ? ????? ??? ????????????? ???? ??? ???????? ?? ?? ?????? ????? ??????? ? ???????? ????? ?????????. ????? ??????? ???? ???????? ? ?????? ????? ? ????????? ?????? ? ????????????? ?????? , ? ?????? ????? ???????? ????????? ??? ?????? ????????? ?????? ???????????????? ??????? ????.
????? ?????? ??????? ?????? ??????? ???????? ? ???????? ???????. ? ????? ??? ???? ????? ???? ????????? ?? ????????????????, ????????, ???????????? ? ????????. ???????????????? ???? ????? ???????? ??? ?????-, ??? ? ????????????????, ? ????? ??????? ?????????? ??????????. ????????? ?? ????? ??? ????????????? ? ??????? ?????????? , ???????? ??? ??????????????????? ????????? ????????? ?? ???????? ?????: ???, ?????? ???? ????????????? ( Hypophthalmichthys molitrix , Hypophthalmichthys nobilis ) ???????? ????????????? ?? ???? ????? ??????? ? ???????? ?????? ????????????? ??????-????????????? ???????????????? ??????????, ??????? ????? ? ????? ????. ??????? ???? ?? ????????? Pomacentridae ???????? ?????????? ????????????????, ?????? ?????? ???? ????? ???????????? ???????, ??? ??? ???????? ??? ?????????, ???????? ??? ????? ?????? ?? ????, ???????? ????????,— ??? ???? ?? ??????? ?? ?? ????? ???????, ? ???? ???????? ???????.
?????? ???? ?????????? ? ???????? ???????? ??????? ?????????? ?????? ????????. ??????????? ??????????? (????? ??? ??????? ????? ) ??????????????? ??????????? ?? ????? ????; ????? ????, ???? ????? ???????? ????????????? , ?????????? , ???????? , ???????? ? ?????????? ??????? , ? ????? ??????? ??????. ???????? ??????????????????? ?? ?????? ??? ???????? ??, ??????? ???????? ?????????????? ?????? ??? ( Labridae , Chaetodontidae ? ??????): ??? ???????? ????????? ? ???????? ??????? ???? ? ??????????? ???? ???-«????????», ??????? ?????????? ???????? ????? ???????? «????????????».
?????? ???? ??? ???????? ??? ??????? ?? ?????????? ?????: ????????, ? ??????? ???????? ???????? ??????????, ? ????? — ?????? ??? ???????? ??????????????? .
????? ? ?????? ????? ???????? ? ???? ??????? ? ??????????????, ? ???????? ???????????? ??? ????????????, ??? ? ??????? ????. ???????????? ???????? ????? ??? ????? ???? ??? ???? (? ??????? ???), ??? ? ???? (? ????????????), ??? ???????????? ???????? ??????? ?????. ???????? ???? — ???????????????, ?? ??? ???? ? ?? ????????? ???????????? ????? ??????? ????, ??? ? ?????????? ?????. ???????????? ????????????? ???????? ??? ???? ??????????? ????????? ??????????? ?????????? ???????? ? ?????????????????? (????) ? ?????. ??????????? ?????? ???????????? ????? ? ????????? ???????? ??? ?????????????? ????????? ????????? ????? ???????, ? ????? ?????????????????? ?????????? ???????, ??????? ??????????? ? ??????????????? ???????. ?????????? ?????? ????????????? ? ??????? ??? ???? ??????, ??? ? ??????-???? ?? ????? ? ?????? ?????????.
????????? ???? ?? ???????? ????????????????. ??????? ????????????? ??????????? ???? , ??????????? ??? ???????????? ?????????? ??????? ??????????, ??????????? ????????? ??????????? ???????????? ?????????? ??????????????? ????????????? ??????? ? ????- ??? ??????????????? ?????.
????? ????????? ??? ?????? ??????? ???? ? ?????????????, ????????? ????????? ????????????? ???????, ??????????? ???????????? ????? ??? ???????? ????? ????? ? ?????? ?? ???? ?????????? Na +-K+-ATPa . ????????? ?????? ?? ???? ?????????? ????? ?????????, ??? ???????????? ? ????????????????????? , ?????? ?????? ???? ? ????????? ?????????????? ????????? ?????????????????.
??????? ????????? ???? ? ?????? (???????????? ?????????? ?????) ????????????? ????? ? ????????? ????????? ?????? ????, ??????? ??? ????????? ????????????? ???????? ??????? ?? ?? ?????? ??????. ??? ?????? ?????????????? ?? ???? ????? ? ?????????? ??????? ????. ??????? ?????? ? ?????? -???? ????????? ?? ????? ????? ????????? ?????? ????????? ????????, ? ?? ????? ??? ??????? ?????? ? ?????????? ?????? ????????? ??????? . ???????????? ???? ???????????? ? ??????????????? ????????? (????????? ? ??? ???????????? ????? ? ????????? ????, ??? ? ?????????? ?????). ???????????? ???????? ? ??? ????????????? ????????? ??????? ????? ?? ?????? ????? ????? ???????? ????????, ? ????? ????????? ????????? ???????? ?????????? ????????.
???? ???, ??????????? ????? ??????? ? ??????? ???????? ????????????? ???????? , ??????????? ???????? ????? ????? ???????? ???????? ?????????? ? ??????????? ?? ????????? ?????: ? ??????????????? ?????? ???????? ????????????, ? ? ?????????????? ????? ??????????????? ?? ?????????? ?????. ???????????????? ???????????, ??? ??????? ??????????????? ????????? ????????? ????, ??? ??????? ???????????, ?????????? ? ???????????? ??????????? ????????????? ??????????? ? ?????????? ????????? ???????? .
? ???????????????? ?????????? ????? ???, ??? ?????????, ???????????? ?????????? ?????? ??????? ???????? , ??????? ???????? ?? ??????? ?? ??????????????? ????? ???????? (?? ???? ??????? ? ??????? ???? ? ???????), ? ????? ????????? , ?????????? ?? ??????? ?? ?????????????? ????? ???????? (?? ??????? ? ???????) .
???????? ??????????? — ???????? ??????? ? ???????????????? ?????. ??? ???? ??????? ??????????? ??? ???? ????? ?????????. ? ????????? ????? ????? ???? ???????? ????????? ??????? ???????? ? ??????????? ??????? ????????? . ?????????????? ????????? ??????? ????????? ?????? ????????? ?????? ???? ??? (? ??????????? ??????? — ?????). ??? ?? ????????? ? ???????? ??????????, ????? ??????????????????? ?? ?????????? ???????? ?????? ? ?? ????????? ?????????? ??????????????? ?????????. ????????? ??????? ???????? ????? ??????????? ? ???? ???????? ? ???????? ????, ?????? ???? (????????, ?????????? ????????), ???????? ???? (????????, ??????? ?? ??????), ???????????? ?????, ???????, ? ????? ??????????? ? ???? ??????? ????? ?????????????, ???????????, ?????????????? ? ??????? ???????????????? ?????? ? ?????. ???????? ??????????? ????? ???? ?????????? , ?????????? ? ?????????????? .
? ???- ????????????? ??? ????? ???????? ? ??????? ?????: ??? ????????????? ?? ??? ???????, ?? ??? ??????? ????? (???????? ??? ???????????????). ???? ??? ????? ????????????????? ????????? ???? — ??????????? ? ?????????? . ???????????????? ???????????? — ??? ?????, ??????? ? ?????? ????? ????? ???????? ???????, ? ??????? ???????????? ???????????? ??????????? ??????? ??????? ? ?????????? ????????? ??????????????? ???????. ????? ????? ?????????????? ???? ?????? ?????????????? ? ????????? ??????? ?????? ( Serranidae ). ??? ?????? ( Labridae ) ???????? ???????????????? ??????????????, ????? ??? ????? ???????? ???????????????? ? ????????? ???????. ? ???? ????????? ?? ????? ???? ????? ?????? ??? ??????? ?????????? ?????, ??? ? ?????????? ????????? ? ?????????. ?????????? ????????? ??????? ??????????? ? ??????? ???????, ??????? ??????? ?? ????? ? ?????? ???????? ?????. ?????? ?????? ??????????????? ?? ???????, ? ?????? ?? ???????? ????????. ???? ?????? ?? ?????? ?????, ?????? ????? (?????? ?? ?????) ????? ???????? ???? ??????????? ?????????, ?????? ????????? ?? ???? ??????? ?????. ???? ?? ?????? ?? ?????? ?????, ????? ??????? ????? ?????? ????????? ?????? ??? ?????, ?????????? ??????? ??????, ??????? ???????????? ?????? ??????. ???? ?? ??? ??????? ? ?????? ?? ?????????? ?????? ?? ??????? ???????????? ???? ????? ? ????????????, ?? ??? ????? ???????? ??????????????? ????????? ?????, ? ????? ????? 14 ???? ?? ??????? ??????? ????????? ??????????, ??????? ????????????? ??????? ??????? ??????.
? ???????? , ??? ??????? ?????????????? ??????????? ?????????? ??????????, ??????? ????????? ???? ?????? ?????????, ? ???? ? ?? ?? ????? ????? ???????? ??? ????????? ??? ?? ?????????? ?????. ? ?????? ???????, ?????????? ??????? (????????, ????????? ????? , ?????? ????? , ??????????? ???????? ), ??? ??????? ?????????????? ?????? ??????????, ?? ?? ?????????? ??????? ?????????? ?????????.
????????? ???????????? ????? ????????????? ??? ??????????, ??? ? ????????????? — ??? ???????????? ????? ??????????? ??????????????????. ???????? ???? ??? ???? ?? ?????? Cyprinodontiformes , ??????? ????????????? ??? ??????????????????? ????????????: ??? ???? ???? ? ??? Rivulus marmoratus . ??? ???? ? Rivulus marmoratus ???????? ?????????? ? ? ?????????? ???????? ? ????????? ???????????? , ??????????? ?????????? ????????. ????? ??????? ????? ?????????? ??????????????? ??????????? ? ????? Hypoplectrus ? Serranus ????????? ???????? ( Percidae ). ???? ??? ???? ???????? ????????????? ????????????? ? ?????????? ????????????, ?? ?????????? ?????? ??????? ??? ????????????? ??? ????????????? ?????? ?????? ????. ???????? ??, ??? ???? ??? ??????? ????? ??????? ????????? ?????, ???? ????? ???? ????? ???????????? ???????? ?????? ? ????????????? ?????????? ?????????? (????) ??? ????????????? (??????).
????????? ????????? ?????? ?????????????? ????????????? , ??????? ? ?????? ???? ? ??? ?? ???????????. ??? ?????????? ???? ????????????? ???????? ? ???????????? ???????, ??????????? ??????? ????????? ???? ? ?????????????? ????????. ? ??????? ?????????? ???????????? ????????? ????????? ??? ???? ???, ?????????????? ? ??????? ?????? ???????. ????? ????????? ????? ??? ???????? ????? ????????????????, ??????? ??? ???????????? ??????????? ?????????? ?????? ?????? ??????, ?????? ??????? ????????????? ?????, ???????? ? ???? ? ?????????? ???????. ??? ???? ???? ?????? ?????? ????? ??????????? ???? ? ?????????????? ??????????????, ??????? ????? ??????????? ?????????? ?????? ????????? ???? ?????-??????????????, ???????? ?????? ?? ???? ????? ????????? ?? ??????? ???????? (??????), ?? ?? ????????? ? ??????? ??????? . ????? ??? ??????????? ????? ???????? ?????????-????????????????. ??? ???? ??? ??????? ????? ????? ?????????? ???????-???????, ??????????? ??????????? ? ??????????? ??????. ???????? ??????????? ???????? ? ????????????? (Cypriniformes): ? ????????? ??????????? ???????????? ????????? ??????????? ?????? Carassius gibelio , ????????? ?????????? ????????-??????????? ???????????? ???? ??????? ???? Cobitis , ????????? ?????????????? ????????????????? (Cyprinodontiformes), ???????? ? ???? Poeciliopsis, ? ????????? ?????? ???.
? ?????????? ? ???? ????? ??????????? ?????????? ??? ??? ?????? ????? ?????? ??????? ????: ???????????? , ???????????????? ? ???????????? .
??? ???????????? ? ???????? ??????? ??????? ? ??????? ?????? ???????????? ? ????, ??? ?????????? ??????????????. ?????? ?????????????? ????? ???? ? ?????????? (??? ? ??????? — Scorpenidae, ? ???????????? ????? — Auchenipteridae ), ?? ????? ????? ????? ?????????? ??????????????? ????. ?? ?????????? ???????? ??????? ?????????? ??? ?????? ????, ??? ? ????????? ? ???? ????? ?????.
??? ????????????????, ???? ??? ????? ????????? ? ????????? ?????, ?????????????? ??????????. ???????? ?? ?????????? ??? ? ???? ?????, ????? ??????????? ?????????? ? ????????? ?? ????????? ????????????? ???????? ??? ???????? ?????? ??????????????. ?????? ? ??? ??????? ?? ????? ?????? ???????? ???????? ???????? ??????????? ???????, ??????? ??????? ? ????. ?????? ??????? ?? ??? ?????? ???????????? ?????????, ????? ???? ?????????? ??????? ??????. ???????? ?????????? ??????, ??????? ??????? ????? ????? ???????????, ???????? ????????????? Poeciliidae — ?????? ???????????????? ? ?????????? ????? ? ????????? . ??? ????? ??????????? ?????????? ????? ??? ????????? .
??????????? ? ????? ???????????? ?? ?????? ??????? ????????????????, ?? ??? ???? ????? ????? ? ??????????? ?????????? ????????? ???????????? ??????? ??? ??????? ????? ?? ??????? ??????????????. ????? ???????, ??????? ??? ???????? ???????? ??????????? ??????????? ???????? ?? ????????? ??????. ???????? ?????? ?????? ????? ??????????? ??? ????, ?? ??? ??????? ????? ? ????????? ????????? ?????, ???????? ??????? ?????? ???????? ?? ????????? ? ?????????????? ????????? Embiotocidae .
????????? ???? ??? ????????? ?????????? ???????? ? ?????????? ??? ??????? ?????. ????? ???? ??? ????? ????? ??????? ??????? , ? ? ??????????? ?? ????, ? ??????????????? ??? ?????????????? ????? ??????????? ????????, ???? ?????????? ???????????? ( ??.-????. ???? «????», ??.-????. ?????? «??????????») ??? ??????????? ( ??.-????. ??? «???????», ??.-????. ?????? «??????????»), ??????????????.
? ???? ???????? ? ?????? ??????? ?????? ?? ????????, ? ?????????? ?????????, ??? ??????? ??????? ? ?????? ????????? ? 10% ?? ?????? ??????? ????? ????????.
???????????, ??? ????? ????????? ????? ???????????? ?????? ? ?? ????, ? ??????? ??? ?????????, ?????? ??????? «????» ???? ???? ?? ?????? .
?? ??????????? ??? ??? ?????? ? ????????? ??????????? ??????????????? ? ????? ? ????????-???????? ????, ? ????? ??????? ? ??????, ? ????? ? ????? ? ??????.
????? ??????????????? ??????? ??????? ????????? ???????????? ?????? ?? ???????? ??????????, ???????? ?? ??????????? ????????. ????? ?? ???????? ? ?????? ?????, ??????? ??????????? ????, ? ????? ???????? ???????? ???? ? ??????.
????? ??????? ????? ?????????? ???????????? ???????? ? ??????? ?????? ? ??????????? ???? ? ????????? ??????? ???????????? ????????????? ????????? . ????? ????????? ?????? ????? ? ?????????, ? ????? ??? ??????????? ????, ???????? ? ???? ????, ???? ???????????????.
?????? ?????????????? ??????? ? ????? ?????????????? ??????????? ??????? . ????? ?????????? ??????? ? ???? ?? ???. ??????? ???????, ????????????? ???????? ??????????? ????????? ? ????? ??????, ?????? ???????????? ???????, ???????????? ?????? ??? ???????? ????????.
??????? ?????????, ????????? ? ??????? ? ?????????, ????????? ? ??????? ??? ? ??????? ??????? .
????????? ? ???????? ???? ???????? ??????? ???????????? ?????? ??????????? ?? ?????.
??? ? ?????? ????????, ???? ????? ?????? ????????????? ?????????????, ??????? ????????????. ??? ?????????????? ???????? ? ??? ???? ????????? ???????? ???????. ??????????????? ?????? ???????? ???? ? ???????, ? ????? ???? ?????, ?????????? ??????????? , ??????? ??????????? ? ????????? ???? ??????????????? . ???? ?????????? ?????????????? ???????? ??? ???????? ?????????, ???????? ???? ????????? ?? ????????????? ?????????????? ????????. ? ????????? ???? ??????? ????? ?????? ?????????????? ? ????????????, ? ????? ? ????????????? ??????, ???????? ??????? ?????? ??????????? ? ????????????? ?????? ? ????? ??????? ??? ? ??? .
????????? ???? ?????????? ???-???????????? ??? ???????? ??????? ?????????. ???????? ?????????? ?? ??? ???????? ??????? ?????? -??????????? ???? Labroides, ????????? ?? ?????????? ????? ? ????????? ? ????? ???????. ????????? ??? ??????? ??????????? ? ???? ????? ???, ??????? ?????????? ?????? ????? ????? ????????? ?????? ? ???? ?? ????, , ????????????, ? ??????? ????? ??????? Etroplus suratensis [ ???? ] .
???????????? ???? ?????????? ?? ??????? ????????????? ??????? ? ?????? ??????? ??? ???????????? ???????? ??????. ????????, ?????????? , ????????? ? ?????????????? ?????? ?????????? ? ???????? ????? (??? ?????????? ) ? ????????? ???????? ????????? (??? ????????? ??????????? ). ??? ?????????? ??????????? ??????? ???? ? ?????. ???????? ???? (????? ? ?????) ???????? ????? ???????? ???????? ????????. ? ??? ???? ??? ?????????????????? ??????, ??????? ????????? ??? ????????????; ???????????? ?????? (?????????? ?????, ??????? ?? ????? ?????????????), ??????? ???????? ??????, ????? ??????? ? ??????? ???????? ? ?????????? ?????? ? ?????????. ? ???? ??????? ?????????? ???????? ???????? ?????? (???????????, ????????? ? ?????????????? ??????). ??? ????? ???????? ???????????????? ??????? ? ?????? ???????? ?????????? (?? ????? ?????? ???????? ?????), ??? ??????????? ? ???????? ????????? ????????? , ?????????????? ?????? ? ?????????. ????????????? ???? (?????????, ?????????? ???? ? ??????) ???????? ???????? ???????? ??? ???????????? ???????????? ? ?????, ??????? ??????? ? ????????? ?????????? (??????????, ??????????? ??????????? ??????? ???????). ?? ?????? ????? ??????? ??????, ?????????? ?????????, ???????????? ?????? ? ????????? ?????????? ?????????? . ??????????????? ????? ???????? ?????? ???????????? ???????, ? ??????? ??????????? ??????????, ???????????, ????????? ? ?????????.
??? ? ? ????????????? ???, ???????? ???????? ????? ????????? ??? (??? ?????????) ???????? ????? (???????? ???????? ?????), ? ??????? ?????????? ????????? ????????? ???????? ??????. ????? ????, ????-???????? ???????? ???????, ?????????? ? ??????????? ????????? ????????? ? ?????? ????????? ???????? (????????, ? ????, ??????, ????????? ? ???????). ??? ? ???????? ??????? ?????????????, ??????????, ?????????? ? ???????????, ??? ????????, ????????? ? ?????????, ? ?? ????? ??? ????????? ???????? ???????? ????? ??????, ?????????????? ? ??????. ? 2006 ???? ????????????? ???????, ??????????? ??????? ? ?????????????, ???? ??????? ? ?????? ???? ????????? ???, ?????-????? . ???? ??? ??? ?? ????????????, ??? ???????, ??-????????, ????? ???, ??? ??????? (?????????????????) ?-?????? ????????????? ? ???????? ??????? ? ????????? .
?- ? ?-?????????, ??????? ??????????????? ? ?-????????? ????????? , ??????????????, ?????????? ? ???? ????????? ???. ?????????????, ?????????? ???????? ??????? ? ????? ???????????????? ? ?????? ???? ????????? ???????????.

? ?????? ?????????????? ??? ????????????? ??? ????? ???? ???????? ???????????, ? ??????? ???????? ??? ????????? ?????? ???????? , ??????? ? ?????? ???. ? ????????? ????? ??????-??????????? ????????????? ??? ??? ???????????????? ?????? , ??? ??? ?????? ????????? ? ??? ???????? ???????????? ??? , ????? ?????????? ?????????????? ??????? ???? ?????? ???????? ??????????? — ???????????? (Tetrapoda), ??????? ?????? ?? ????????.
??????????? ??? ????????? ?? 3 ??????: ????????, ????????? ? ???????????? ????. ??? ?? ?????? ???????? XX ???? ????????? ??? ?????? ? ???????? ?????????? ?????????? ? ????? ??????? ???, ?????? ? ????????? ??????????? ???????????? ?????????? ??????????????? ??? ??????????????? ??????. ??? ?? ?????, ????????? ???????????? ????????????? ? ????????? ???? ??????? ????? ?????????? ????????? ? ???????????? ??? ? ???? ????? Osteichthyes .
????????? ???????? ???????? ??? ???????????? ????:
???? ????? ? ????, ??????? ???????? ???????? ????????????. ????? 361 ??? ??², ??? 71 % ???? ??????????? ??????? ????, ?????? ?????? ? ????????, ? 2,5 ??? ??² — ??????????? ?????????. ???? ?????????????? ?? ???????????? ???????? (????? 6000 ? ??? ??????? ??????) ?? ?????????? ?????? ???????? ?????? (11022 ?). ???? ??????????? ????? ? ???????? ????? ? ??????????? ?????.
? ????????? ????? ???? — ?????????????? ?????? ???????? ? ?????? ?????????? . ?????? ? ?????????????, ??? ????????? ???? ???????.
???? ?????????????? ? ????????? ???????? ?????? ?????. ???????, ???????? ?? ???, ??????? ??:
? ???????? ?????? ???????????? ???????? ????????? ??????????? ????, ?????????, ?????????? ????? ? ??.
?? ?????? ???????? ????????? ???????, ???????????? ? ????????? ???.
??? ?????????? ??????? ???????????? ????? ?????? ????? ???? ??? ????? ???????? ? ?????? ?????????? ????????:
?????? ???? ??? ?????????? ? ?????? — ? ?????????? . ?????????? ? ?????????? ??????????? ??? ? ????????? ??????? ???????? ????????? ?????? ?? ????, ?????????? ? ???????? ?????? ????????.
? ????????????? ????????? ???????????? ???? ?????? ?????????? ???????? , ?????????? ??? ??? ???????? ???????????? ? ???????? ???????????? ????????????? ??????.
? 1961 ???? ? ??????? ???????? ?. ???????? ??? ??????? ?????????????? ????? ???? ??? ?????? ???????????? ????????? ??????? ? ???????????, ?????????? ?????????? , ??? ? ???????????? ??????? ????????? ???????????? ??????????? ?? ????.
????????????? ???????????? ???????? ???????? ????????? ??? ?????? ??????????? ????????? ??? ??? ???? ????? ???????? ??? ?????????????, ????? ??????????? ???, ???????????????? ????? ?????, ????????? ????? ??? ? ????????? ???????.
????, ??? ? ???? , ? ????????? ??????????? ??????, ?????? ? ??????? ????????? ?????? ???? ???? ? ????????? ? ??????? ????????? ??????? ????.