Австралопитеки
- 1 year ago
- 0
- 0

Австралопите́ки ( лат. Australopithecus от лат. «южный» + др.-греч. «обезьяна») — род ископаемых высших приматов , обладавших признаками прямохождения и антропоидными чертами в строении черепа, чей хронологический период (как рода) определяется от 4,2 до 1,8 млн лет назад. Также понятие «австралопитеки» часто используется шире и распространяется на большую эволюционную группу гоминидов — австралопитецина (или хоминина) , включающую в себя, помимо рода Australopithecus , представителей ещё пяти родов: Sahelanthropus , Orrorin , Ardipithecus , Kenyanthropus и Paranthropus , либо вместе с парантропами выделяют в подтрибу (англ.) (в трибе гоминини ( Hominini ) .
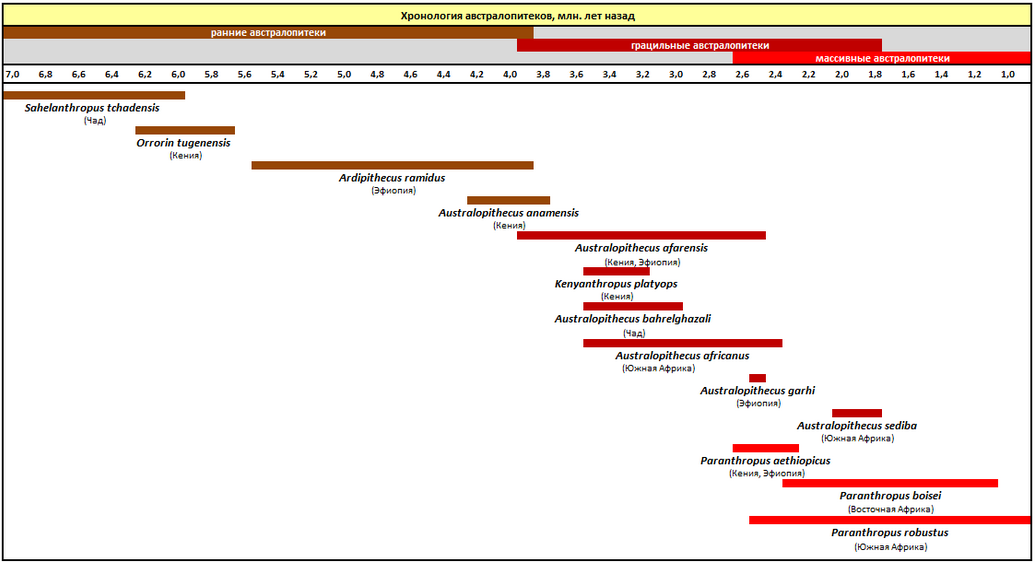
Кости австралопитеков впервые были обнаружены Раймондом Дартом в карьере каменоломни Таунг ( Трансвааль ) в 1924 году, а затем — в Восточной ( Олдувай ) и Центральной Африке . Являются вероятными предками рода Люди ( Homo ) . Судя по данным палеонтологии , род австралопитеков развивался в Восточной Африке начиная с 4 млн лет назад, впоследствии эволюционировав, он распространился по всей Африке и вымер 2 млн лет назад. За это время появилось несколько видов австралопитеков, в том числе Australopithecus afarensis , A. africanus , A. anamensis , A. bahrelghazali , A. deyiremeda (предлагается), A. garhi и A. sediba . В германском Эппельсхайме найдены зубы европейских гоминидов возрастом почти 10 млн лет — Epp 13.16 и Epp 30.16 (верхний левый клык и верхний правый первый моляр ), схожие с зубами африканских австралопитеков .
Австралопитеки сыграли значительную роль в эволюции человека. Вид Homo sapiens , или Человек разумный через промежуточные виды ведёт своё происхождение от австралопитеков. Предки человека отделились от общего с австралопитеками корня примерно 3 млн лет назад. Также предполагается, что австралопитеки были первыми гоминидами, обладавшими набором специфичных белков, кодирующихся геном ( дупликация 3,4 млн л. н. называется SRGAP2B, дупликация 2,4 млн л. н. — , дупликация ~1 млн лет назад — SRGAP2D). Эти гены отвечают за увеличение длины и активность нейронов в мозге . От одного из видов австралопитека в итоге и произошёл род Homo около 2 млн л. н. Примером может служить Homo habilis — человек умелый. В процессе эволюции появились современные люди — H. sapiens sapiens — человек современной анатомии , или неоантроп .
Из-за сложностей морфологического разделения внутри семейства Hominidae , а также для лучшего понимания эволюционного развития гоминид учёные выделяют большую группу ископаемых приматов — австралопитековые , или австралопитеки, куда, помимо собственно рода Australopithecus, входят и другие роды. Вследствие этого в литературе австралопитеки могут рассматриваться как в узком (род), так и широком смысле слова (эволюционная группа). В этом контексте современная палеоантропология условно разделяет австралопитеков на три группы:
Ранние австралопитеки включают в себя роды:
Грацильные австралопитеки включают в себя следующие виды:
Последняя группа из-за своей специфической анатомии выделяется в отдельный род — Paranthropus , насчитывающий три вида:
Существует ещё несколько спорных видов, которые могут быть отнесены к группе австралопитеков, однако это выходит за рамки настоящей статьи.

Австралопитеки жили в плиоцене , примерно от 4,2 млн лет назад — Australopithecus anamensis ( ( ) — 3,67 млн лет ) до менее миллиона лет назад. На временно́й шкале хорошо прослеживаются три длинных эпохи основных видов, примерно по миллиону лет на вид. Большинство видов австралопитеков было всеядно , однако были подвиды, специализировавшиеся на растительной пище. Предком основного вида, скорее всего, был вид A. anamensis , а первым основным видом, известным на данный момент, стал вид A. afarensis , который просуществовал примерно 1 млн л. н. По-видимому, морфологически эти существа были не более, чем обезьянами, по-человечески перемещавшимися на двух ногах, хотя и сгорбленно. Возможно, под конец они умели пользоваться подручными камнями для раскалывания, к примеру, орехов.
Считается, что A. afarensis в конце разделился на два подвида: первая ветвь пошла к очеловечиванию и Homo habilis , вторая продолжила совершенствоваться в австралопитеках, образовав новый вид A. africanus . У A. africanus были немного менее развиты конечности, чем у A. afarensis , но зато они научились пользоваться подручными камнями, палками и острыми осколками костей и уже могли самостоятельно изготавливать простейшие костяные и деревянные орудия. Через миллион лет образовали два новых высших и последних известных подвида австралопитеков A. boisei и A. robustus , которые просуществовали вплоть до 900 тыс. лет до н. э.
С точки зрения таксономии австралопитеков относят к семейству гоминидов (включающему также людей и современных крупных человекообразных обезьян ). Вопрос о том, были ли какие-либо австралопитеки предками людей, или они представляют собой «сестринскую» по отношению к людям группу, не выяснен до конца.
Грацильные австралопитеки передали современным обезьянам и людям ряд черт — они были широко распространены в Восточной и Северной Африке 3,5 млн лет назад. Самые ранние свидетельства ходивших постоянно на двух ногах гоминидов найдены в Танзании — Лаэтоли . Этот археологический объект содержит ряд находок следов, оставшихся от гоминид, которые очень похожи на следы современных людей, их возраст оценивается в 3,6 млн лет. Следы единогласно отнесены именно к австралопитекам, поскольку в тот период они населяли те места .
На данный момент вид австралопитеков, от которого отделился предок рода Homo , не известен. Изначально предполагалось, что это A. africanus , но в дальнейшем находки, имеющие явное отношение к роду Homo , показали дальнейшее увеличение возраста и стали старше A. africanus . В итоге учёные считают, род Homo отделился либо от вида A. afarensis , либо от Kenyanthropus . При этом не исключается, что общий предок рода Homo и этих видов жил ещё раньше. Согласно исследованиям генетиков из проекта Chimpanzee genome project, генетические линии Ardipithecus , Australopithecus и Homo и шимпанзе Pan troglodytes и Pan paniscus разошлись от общего предка 5—6 млн лет назад .
Сахелантроп имеет древность 7 млн лет, Оррорин — около 6 млн лет. Эти виды жили до того, как генетические линии рода люди и шимпанзе окончательно разошлись, по меньшей мере на 1 млн лет позднее, согласно молекулярным часам .
Австралопитеки жили в саваннах, популяции были с упором на большее количество самцов. Самцы сотрудничали, с целью добычи пропитания, охраны района проживания группы и защиты самок и детёнышей. Самцы спаривались с несколькими самками, практикуя своеобразный вариант многожёнства. Когда детёныши вырастали и достигали полового созревания , самцы оставались в группе, а самки уходили в другие места, искать другие группы для жизни. В данный исторический период точно так же поступают самки шимпанзе в Африке .
Не существует никаких доказательств использования австралопитеками естественных убежищ или создания ими временных наземных укрытий для ночлега из веток или иных материалов. Однако, не вызывает сомнений, что они могли, подобно современным шимпанзе, ночевать на деревьях, сооружая там временные гнёзда .
Продолжительность жизни австралопитеков, которая постоянно подвергалась угрозе не только со стороны хищников, но и некоторых крупных травоядных, была незначительной и в среднем составляла 20 лет. Практически никто из них не доживал до 40 лет, и лишь один из семи доживал до 30 .

C человеком австралопитеков сближает слабое развитие челюстей, отсутствие крупных выступающих клыков, хватательная кисть с развитым большим пальцем, опорная стопа и строение таза, приспособленное для прямохождения. Головной мозг относительно крупный (530 см³), но по строению мало отличающийся от мозга современных человекообразных обезьян . По объёму он составлял не более 35 % от средних размеров мозга современного человека. Размеры тела также были невелики, не более 120—140 см в высоту, телосложение стройное. Предполагается, что разница в размерах самцов и самок австралопитеков была больше, чем у современных гоминидов.
Например, у современных людей мужчины в среднем лишь на 15 % крупнее женщин, в то время как у австралопитеков они могли быть на 50 % выше и тяжелее , что порождает дискуссии о принципиальной возможности столь сильного полового диморфизма у этого рода гоминидов. Одним из основных характерных признаков для парантропов является костяной стреловидный гребень на черепе, присущий самцам современных горилл, поэтому нельзя до конца исключить, что робустерные/ парантропные формы австралопитеков являются самцами, а грацильные — самками, альтернативным объяснением может быть отнесение форм разных размеров к разным видам или подвидам.
Согласно мнению исследователей, пропорции тела австралопитеков сближают их с бонобо. Один из ведущих эволюционных биологов Джереми Гриффит предположил, что бонобо может быть фенотипически подобен австралопитекам. Так же, согласно моделям терморегуляции , австралопитеки были полностью покрыты волосами, как и шимпанзе и бонобо, что заметно отличает их от людей.
Основным кандидатом на место предка австралопитеков является род Ardipithecus . При этом самый древний из представителей нового рода, Australopithecus anamensis , произошёл непосредственно от Ardipithecus ramidus 4,4—4,1 миллиона лет назад, а 3,6 миллиона лет назад дал начало Australopithecus afarensis , к которому принадлежит первая найденная современными людьми — « Люси ». С находкой в 1985 году так называемого «чёрного черепа», который был очень похож на Paranthropus boisei , с характерным костяным гребнем, но при этом был на 2,5 млн старше.
В родословной австралопитеков появилась официальная неопределённость, поскольку, хоть и результаты анализов могут сильно меняться в зависимости от множества обстоятельств и среды, где находился череп , и, как водится, будут ещё десятилетиями перепроверяться десятки раз, но на текущий момент получается, что Paranthropus boisei не мог произойти от Australopithecus africanus , поскольку жил до них, и, как минимум, жил в одно время с Australopithecus afarensis , и, соответственно, тоже не мог от них произойти, если, конечно, не учитывать гипотезу, что парантропные формы австралопитеков и австралопитеки являются самцами и самками одного и того же вида.
Род Australopithecus считается предком по меньшей мере двух групп гоминидов: парантропов и людей , хотя по уровню интеллекта австралопитеки мало отличались от обезьян. Их главным отличием от обезьян до них стало прямохождение, в то время как большинство обезьян является четвероногим. Таким образом, прямохождение предшествовало развитию интеллекта у людей, а не наоборот, как ранее предполагалось.
Как австралопитеки перешли к прямохождению, пока не ясно. Среди рассматриваемых причин называют необходимость хватать объекты, такие как пища и детёныши, передними лапами, и осматривать окрестности поверх высокой травы в поисках пищи или чтобы вовремя заметить опасность. Высказывается также предположение , что общие предки прямоходящих гоминидов (в том числе людей и австралопитеков) жили на мелководье и питались мелкими водными обитателями, а прямохождение сформировалось как адаптация к передвижению по мелководью.
Согласно данным генетики, признаки прямохождения появились у некоторых вымерших видов обезьян ещё около 6 млн лет назад, в эпоху расхождения между людьми и шимпанзе . Это значит, что не только сами австралопитеки, но и вид, который был их предком, например, ардипитек , уже мог быть прямоходящим. Возможно, прямохождение было элементом адаптации к жизни на деревьях. Современные орангутаны используют все четыре лапы для перемещения только по толстым сучьям, в то время как к более тонким ветвям либо цепляются снизу, либо идут по ним на задних лапах, готовясь передними уцепиться за другие ветви повыше или балансируя для устойчивости.
Такая тактика позволяет им приблизиться к плодам, расположенным далеко от ствола, или прыгать с одного дерева на другое. Изменения климата, произошедшие 11—12 млн лет назад, привели к сокращению лесных массивов в Африке и появлению больших открытых пространств, что и могло подтолкнуть предков австралопитеков к переходу к прямохождению по земле. В отличие от них, предки современных шимпанзе и горилл специализировались на лазании по вертикальным стволам и лианам, чем обусловлена их кривоногая и косолапая походка на земле. Тем не менее, люди унаследовали много общего с этими обезьянами, включая строение костей рук, усиленных для хождения с опорой на костяшки пальцев .

Не исключено также, что австралопитеки не были прямыми предками людей, а представляли собой тупиковую ветвь эволюции. К таким выводам подталкивают, в частности, недавние находки сахелантропа , ещё более древней человекообразной обезьяны, которая была больше похожа на Homo erectus , чем австралопитеки. В 2008 году был открыт новый вид австралопитеков, A. sediba , живший в Африке менее двух миллионов лет назад. Хотя по отдельным морфологическим признакам он ближе к людям, чем более древние виды австралопитеков, что дало основание его открывателям объявить его переходной формой от австралопитеков к людям, одновременно с ним, по-видимому, уже существовали первые представители рода Homo , такие, как человек рудольфский , что исключает возможность того, что этот вид австралопитеков мог быть предком современного человека.
Большинство видов австралопитеков использовало орудия труда не более современных обезьян. Известно, что шимпанзе и гориллы способны колоть орехи камнями, использовать палочки для извлечения термитов и дубинки для охоты. Насколько часто австралопитеки охотились — вопрос спорный, так как их ископаемые остатки редко ассоциируют с остатками убитых животных.

Возникновение бипедализма среди австралопитеков приписывается африканскому австралопитеку , по предположениям учёных, именно его следы были найдены в месте Лаэтоли , Танзания. Таким образом, бипедализм скорее всего появился намного раньше, чем крупный мозг, резко отделивший род Хомо от остальных ветвей обезьян. Бипедальность является главным шагом, позволившим освободить руки от постоянной нагрузки при ходьбе и беге. После этого эволюционного шага руки гоминид освободились, стало возможно держать предметы при ходьбе, а не только сидя, как это могут делать современные шимпанзе, но им намного сложнее передвигаться и одновременно манипулировать предметами.
Так же бипедальность позволила постоянно носить матери ребёнка на руках. Согласно исследованию вида Orrorin tugenensis — Оррорин, жившего 6 млн лет назад, он также имел способность использовать бипедальность. Данные сведения привели учёных к теории, что бипедальность возникла как адаптация к передвижению по ветвям и уже затем перешла к роду Хомо как способ передвижения по земле.
Современные орангутаны на Суматре используют четыре ноги при ходьбе по толстым ветвям, но, когда толщина ветвей снижается до 4 см, они встают на задние ноги и передвигаются бипедально, пользуясь руками как поддержкой при ходьбе. Такой механизм передвижения даёт орангутанам возможность подбираться к плодам на самом краю ветвей и обеспечивает больший запас еды в сложные периоды . Предполагается, что предки горилл и шимпанзе приобрели подвижное, согнутое бедро и сгибающееся колено как механизм для того, чтобы взбираться по стволам деревьев наверх. Подобная адаптация вызвана изменением климата 12 — 11 млн лет назад, когда леса Восточной и Центральной Африки увеличились по площади, и передвигаться по земле, которую стал покрывать полог из растительности, стало не так удобно, как раньше. Люди имеют много общих черт именно с этими видами обезьян — в том числе и запястья, которые значительно усилились для больших нагрузок.


По современным реконструкциям учёные предполагают, что, возможно, имеет место пример конвергентной эволюции, когда роды обезьян вырабатывали схожие признаки, не являясь ближайшими родственникам, а были отдельно развивающимися ветвями после отделения от общего предка . Радикальные изменения в строении тела произошли ещё до грацильных австралопитеков, а значит — раньше 4 млн лет назад . В результате находок древностью 3 — 4 млн лет в 2010 году было выяснено, что австралопитеки умели пользоваться примитивными ножами из камня, чтобы вырезать куски мяса из туши .
Количество зубов у австралопитеков, как у людей — 32. У них были такие же небольшие клыки и это делает их отличными от предыдущих гоминидов . Эмаль на зубах была такой же толщины, как и у рода Хомо . Это отличает зубы австралопитеков от других больших обезьян. Объяснением такому приспосабливанию служит то, что эти гоминины проводили на земле больше времени, чем на деревьях, и питались в большом количестве клубнями , орехами и зёрнами злаков . В силу того, что гоминины в тот период ещё не научились мыть эти растения перед едой, песок и грязь на них наносили дополнительные повреждения эмали зубов и адаптация вызвала её утолщение. Грацильные австралопитеки, судя по зубам, включали в свой рацион значительное количество мясной пищи .
A. afarensis , судя по состоянию зубов, употреблял много фруктов и листьев, и мало — трав и семян. A. anamensis напротив, активно ел травы и семена, дополняя ими рацион из фруктов и листьев .
Судя по количеству стабильных изотопов кальция в эмали зубов, период грудного вскармливания у ранних Homo длился три-четыре года, тогда как у африканских австралопитеков и парантропов ( Paranthropus robustus ) период грудного вскармливания длился всего несколько месяцев .
Австралопитеки питались фруктами, овощами, небольшими ящерицами и клубнями. Вид P. robustus питался орехами и семенами, но основной была не твёрдая, а более мягкая еда. В 2005 году в Эфиопии в местности Гона были найдены кости со следами разделки орудиями, древностью 2,6 млн лет. Ископаемые данные говорят о том, что между 2,6 — 2,5 млн лет назад потребляли мясо минимум три вида гомининов — A. africanus , A. garhi и P. aethiopicus . По мнению ряда учёных, австралопитеки не брезговали и падалью , подбирая её за хищниками.

В период между 9 и 5 миллионами лет назад находки австралопитеков почти отсутствуют. Однако были совершены находки большого количества обезьян возрастом от 15 до 10 миллионов лет, в том числе вида . Затем, начиная с 4 миллионов лет назад, находки также имеются в большом количестве. Подобная странность породила теорию о том, что в период среднего миоцена произошли некие серьёзные колебания климата, растительности и рельефа, которые послужили причиной такому странному провалу в количестве и разнообразии видов австралопитеков и других обезьян.
15 миллионов лет назад Африка была покрыта лесами от Атлантического до Индийского океана. 12 млн лет назад произошли тектонические изменения поверхности, дрейф литосферных плит на севере от Восточной Африки привёл к изменению рельефа в Восточно-Африканская рифтовой долине и она заблокировала поступление влажных воздушных масс в Восточную Африку. Это вызвало разрушение прежнего почти сплошного лесного покрова в этой части континента, его фрагментирование. Деревья большей частью высохли, сохранились лишь островками, как в современной Африканской саванне, также сохранились кустарники, они в тот период препятствовали образованию излишне больших открытых пространств, какие есть в современной Африке. В дальнейшем, начиная с 3 миллионов лет назад, появился современный тип саванн, и постоянные миграции крупных стад животных в этих саваннах.

В результате стабильная и однородная среда распалась и стала более разнообразной по условиям для жизни. Популяции гоминин, до этого свободно друг с другом пересекавшиеся в сплошном лесу, оказались разделены в разных условиях. Лесные участки были более прохладными по температуре, ландшафты без деревьев стали намного более жаркими.
Анализ современного состояния климата в Африке и его сравнение с данными палеоклиматологии, вкупе с хронологией найденных гоминидов, вывели следующую теорию:
Бахр-эль-газальский австралопитек был найден в Чаде французским палеонтологом Мишелем Бруне в местности Коро Торо на территории древнего речного русла Бахр-эль-Газаль на юге административного региона Борку , к западу от Рифтовой долины. А значит, если принять данный вид за самостоятельный, то теория имеет серьёзный изъян. Однако, в сообществе учёных нет единого мнения, и данный вид австралопитека может быть подвидом Афарского австралопитека .
Также обнаружены остатки предположительно бипедальных гоминидов, не связанных с саваннами — Оррорин , Сахелантроп , Ардипитеки .