
Бизнес-экосистема
- 1 year ago
- 0
- 0


Экосисте́ма , или экологи́ческая систе́ма (от др.-греч. οἶκος — жилище, местопребывание и σύστημα — «целое, составленное из частей; соединение») — основная природная единица на поверхности Земли , совокупность совместно обитающих организмов (биотических) и условий их существования (абиотических), находящихся в закономерной взаимосвязи друг с другом и образующих систему .
Одно из основных понятий в науке Экология . Это биологическая система , состоящая из сообщества живых организмов ( биоценоз ), среды их обитания ( биотоп ), системы связей, осуществляющей обмен веществ и энергии между ними.
Пример экосистемы — пруд с обитающими в нём растениями , рыбами , беспозвоночными животными , микроорганизмами , составляющими живой компонент системы, биоценоз. Для пруда как экосистемы характерны донные отложения определённого состава, химический состав ( ионный состав, концентрация растворённых газов ) и физические параметры ( прозрачность воды , сезонный ход температуры), а также определённые показатели биологической продуктивности , водоёма и специфические условия данного водоёма. Другой пример экологической системы — лиственный лес в средней полосе России с определённым составом лесной подстилки, характерной для этого типа лесов почвой и устойчивым растительным сообществом , и, как следствие, со строго определёнными показателями микроклимата (температуры, влажности , освещённости ) и соответствующим таким условиям среды комплексом животных организмов . Немаловажным аспектом, позволяющим определять типы и границы экосистем, является трофическая структура сообщества и соотношение производителей биомассы , её потребителей и разрушающих биомассу организмов , а также показатели продуктивности и обмена вещества и энергии . [ источник не указан 908 дней ]
Идеи единства всего живого в природе, его взаимодействия и обусловливания процессов в природе ведут своё начало с античных времён. Однако, приобретать современную трактовку понятие стало в конце XIX века:
В 1935 году современный термин экосистема впервые был предложен английским экологом Артуром Тенсли .
В смежных науках существуют также различные определения, в той или иной степени отражающие понятие «экосистема», например:


Иногда особо подчёркивается, что экосистема — это исторически сложившаяся система (см. ).
Экосистема — сложная (по определению сложных систем Л. Берталанфи ) самоорганизующаяся, саморегулирующаяся и саморазвивающаяся система. Основной характеристикой экосистемы является наличие относительно замкнутых, стабильных в пространстве и времени потоков вещества и энергии между биотической и абиотической частями экосистемы . Из этого следует, что не всякая биологическая система может назваться экосистемой, например, таковыми не являются аквариум или трухлявый пень . Данные биологические системы (естественные или искусственные) не являются в достаточной степени самодостаточными и саморегулируемыми (аквариум), если перестать регулировать условия и поддерживать характеристики на одном уровне, достаточно быстро она разрушится. Такие сообщества не формируют самостоятельных замкнутых циклов вещества и энергии (пень), а являются лишь частью большей системы . Такие системы следует называть сообществами более низкого ранга, или же микрокосмами . Иногда для них употребляют понятие — фация (например, в геоэкологии ), но оно не способно в полной мере описать такие системы, особенно искусственного происхождения. В общем случае в разных науках понятию «фация» соответствуют различные определения: от систем субэкосистемного уровня (в ботанике, ландшафтоведении) до понятий, не связанных с экосистемой (в геологии), либо понятие, объединяющее однородные экосистемы ( Сочава В. Б. ), или почти тождественное ( Берг Л. С. , Раменский Л. Г. ) определению экосистемы.

Экосистема является открытой системой и характеризуется входными и выходными потоками вещества и энергии . Основа существования практически любой экосистемы — поток энергии солнечного света , который является следствием термоядерной реакции, — в прямом (фотосинтез) или косвенном (разложение органического вещества) виде, за исключением глубоководных экосистем: « чёрных » и « » курильщиков, источником энергии в которых является внутреннее тепло Земли и энергия химических реакций .
В соответствии с определениями, между понятиями «экосистема» и «биогеоценоз» нет никакой разницы, биогеоценоз можно считать полным синонимом термина экосистема . Однако существует распространённое мнение, согласно которому биогеоценоз может служить аналогом экосистемы на самом начальном уровне , так как термин «биогеоценоз» делает бо́льший акцент на связь биоценоза с конкретным участком суши или водной среды, в то время как экосистема предполагает любой абстрактный участок. Поэтому биогеоценозы обычно считаются частным случаем экосистемы . Разными авторами в определении термина биогеоценоз перечисляются конкретные биотические и абиотические компоненты биогеоценоза, в то время как определение экосистемы носит более общий характер .
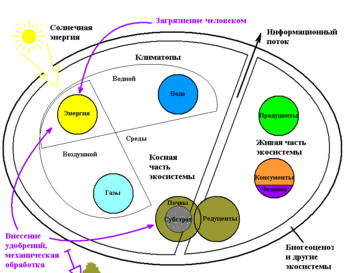
В экосистеме можно выделить два компонента — биотический и абиотический. Биотический делится на автотрофный (организмы, получающие первичную энергию для существования из фото- и хемосинтеза или продуценты) и гетеротрофный (организмы, получающие энергию из процессов окисления органического вещества — консументы и редуценты) компоненты , формирующие трофическую структуру экосистемы.
Единственным источником энергии для существования экосистемы и поддержания в ней различных процессов являются продуценты , усваивающие энергию солнца , ( тепла , химических связей ) с эффективностью 0,1—1 %, редко 3—4,5 % от первоначального количества. Автотрофы представляют первый трофический уровень экосистемы. Последующие трофические уровни экосистемы формируются за счёт консументов (2-й, 3-й, 4-й и последующие уровни) и замыкаются редуцентами , которые переводят неживое органическое вещество в минеральную форму (абиотический компонент), которая может быть усвоена автотрофным элементом .
С точки зрения структуры в экосистеме выделяют :
Последние три компонента формируют биомассу экосистемы.
С точки зрения функционирования экосистемы выделяют следующие функциональные блоки организмов (помимо автотрофов):
Данное разделение показывает временно-функциональную связь в экосистеме, фокусируясь на разделении во времени образования органического вещества и перераспределении его внутри экосистемы (биофаги) и переработки сапрофагами . Между отмиранием органического вещества и повторным включением его составляющих в круговорот вещества в экосистеме может пройти существенный промежуток времени, например, в случае соснового бревна, 100 и более лет.
Все эти компоненты взаимосвязаны в пространстве и времени и образуют единую структурно-функциональную систему.

Обычно понятие экотоп определялось как местообитание организмов, характеризующееся определённым сочетанием экологических условий: почв, грунтов, микроклимата и др. В этом случае это понятие близко к понятию климатоп .
На данный момент под экотопом в отличие от биотопа понимается определённая территория или акватория со всем набором и особенностями почв , грунтов , микроклимата и других факторов в неизменённом организмами виде . Примерами экотопа могут служить наносные грунты, новообразовавшиеся вулканические или коралловые острова , вырытые человеком карьеры и другие заново образовавшиеся территории. В этом случае климатоп является частью экотопа.

Изначально «климатоп» был определён В. Н. Сукачёвым (1964) как воздушная часть биогеоценоза, отличающаяся от окружающей атмосферы своим газовым составом, особенно концентрацией углекислого газа в приземном биогоризонте, кислорода там же и в биогоризонтах фотосинтеза , воздушным режимом, насыщенностью , уменьшенной и изменённой солнечной радиацией и освещённостью, наличием люминесценции растений и некоторых животных, особым тепловым режимом и режимом влажности воздуха .
На данный момент это понятие трактуется чуть более широко: как характеристика биогеоценоза, сочетание физических и химических характеристик воздушной или водной среды, существенных для населяющих эту среду организмов . Климатоп задаёт в долговременном масштабе основные физические характеристики существования животных и растений, определяя круг организмов, которые могут существовать в данной экосистеме.
Под эдафотопом обычно понимается почва как составной элемент экотопа . Однако более точно это понятие следует определять как часть косной среды, преобразованной организмами, то есть не всю почву , а лишь её часть . Почва (эдафотоп) является важнейшей составляющей экосистемы: в нём происходит замыкание циклов вещества и энергии, осуществляется перевод из мёртвого органического вещества в минеральные и их вовлечение в живую биомассу . Основными носителями энергии в эдафотопе выступают органические соединения углерода , их лабильные и стабильные формы, они в наибольшей степени определяют плодородие почв.

« Биотоп » — преобразованный биотой экотоп или, более точно, участок территории, однородный по условиям жизни для определённых видов растений или животных, или же для формирования определённого биоценоза .
Биоценоз — исторически сложившаяся совокупность растений, животных, микроорганизмов, населяющих участок суши или водоёма (биотоп). Не последнюю роль в формировании биоценоза играет конкуренция и естественный отбор . Основная единица биоценоза — консорция , так как любые организмы в той или иной степени связаны с автотрофами и образуют сложную систему консортов различного порядка, причём это сеть является консортом всё большего порядка и может косвенно зависеть от всё большего числа детерминантов консорций.
Также возможно разделение биоценоза на фитоценоз и зооценоз . Фитоценоз — это совокупность растительных популяций одного сообщества, которые и формируют детерминантов консорций. Зооценоз — это совокупность популяций животных, которые и являются консортами различного порядка и служат механизмом перераспределения вещества и энергии внутри экосистемы (см. функционирование экосистем).
Биотоп и биоценоз вместе формируют биогеоценоз/экосистему.

Экосистема может быть описана комплексной схемой положительных и отрицательных обратных связей , поддерживающих гомеостаз системы в некоторых пределах параметров окружающей среды . Таким образом, в некоторых пределах экосистема способна при внешних воздействиях поддерживать свою структуру и функции относительно неизменными. Обычно выделяют два типа гомеостаза: резистентный — способность экосистем сохранять структуру и функции при негативном внешнем воздействии (см. Принцип Ле Шателье — Брауна ) и упругий — способность экосистемы восстанавливать структуру и функции при утрате части компонентов экосистемы . В англоязычной литературе используются сходные понятия: локальная устойчивость — англ. local stability (резистентный гомеостаз) и общая устойчивость — англ. global stability (упругий гомеостаз) .

Иногда выделяют третий аспект устойчивости — устойчивость экосистемы по отношению к изменениям характеристик среды и изменению своих внутренних характеристик . В случае, если экосистема устойчиво функционирует в широком диапазоне параметров окружающей среды и/или в экосистеме присутствует большое число взаимозаменяемых видов (то есть, когда различные виды, сходные по экологическим функциям в экосистеме, могут замещать друг друга), такое сообщество называют динамически прочным (устойчивым). В обратном случае, когда экосистема может существовать в весьма ограниченном наборе параметров окружающей среды, и/или большинство видов незаменимы в своих функциях, такое сообщество называется динамически хрупким (неустойчивым) . Данная характеристика в общем случае не зависит от числа видов и сложности сообществ. Классическим примером может служить Большой Барьерный риф у берегов Австралии (северо-восточное побережье), являющийся одной из «горячих точек» биоразнообразия в мире — симбиотические водоросли кораллов, динофлагелляты , весьма чувствительны к температуре. Отклонение от оптимума буквально на пару градусов ведёт к гибели водорослей, а до 50—60 % (по некоторым источникам до 90 %) питательных веществ полипы получают от фотосинтеза своих мутуалистов .
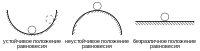
У экосистем существует множество состояний, в которых она находится в динамическом равновесии; в случае выведения из него внешними силами, экосистема совершенно необязательно вернётся в изначальное состояние, зачастую её привлечёт ближайшее равновесное состояние ( аттрактор ), хотя оно может быть очень близким к первоначальному .

Обычно устойчивость связывали и связывают с биоразнообразием видов в экосистеме ( альфаразнообразие ), то есть, чем выше биоразнообразие, чем сложнее организация сообществ, тем сложнее пищевые сети, тем выше устойчивость экосистем. Но уже 40 и более лет назад на данный вопрос существовали различные точки зрения, и на данный момент наиболее распространено мнение, что как локальная, так и общая устойчивость экосистемы зависят от значительно большего набора факторов, чем просто сложность сообществ и биоразнообразие. Так, на данный момент с повышением биоразнообразия обычно связывают повышение сложности, силы связей между компонентами экосистемы, стабильность потоков вещества и энергии между компонентами .

Важность биоразнообразия состоит в том, что оно позволяет формировать множество сообществ, различных по структуре, форме, функциям, и обеспечивает устойчивую возможность их формирования. Чем выше биоразнообразие, тем большее число сообществ может существовать, тем большее число разнообразных реакций (с точки зрения биогеохимии ) может осуществляться, обеспечивая существование биосферы в целом .
На данный момент не существует удовлетворительного определения и модели, описывающей сложность систем и экосистем в частности. Существует два широко распространённых определения сложности: колмогоровская сложность — слишком специализированное для применения к экосистемам. И более абстрактное, но тоже неудовлетворительное определение сложности, данное И. Пригожиным в работе «Время, хаос, квант» : Сложные системы — не допускающие грубого или операционального описания в терминах детерминистских причинностей . В других своих трудах И. Пригожин писал, что не готов дать строгого определения сложности , поскольку сложное — это нечто, что на данный момент не может быть корректно определено.
В качестве параметров сложности экосистем традиционно подразумевались общее число видов (альфа-разнообразие), большое число взаимодействий между видами, сила взаимодействий между популяциями и различные сочетания этих характеристик. При дальнейшем развитии этих представлений появилось утверждение, что чем больше путей переноса и преобразования энергии в экосистеме, тем она устойчивей при различных видах нарушений .
Однако, позже было показано, что данные представления не могут охарактеризовать устойчивость экосистем . Существует множество примеров как весьма устойчивых монокультурных сообществ (фитоценозы орляка), так и слабоустойчивых сообществ с высоким биоразнообразием (коралловые рифы, тропические леса). В 70-80-х годах XX столетия усилился интерес к моделированию зависимости устойчивости от сложности экосистем . Разработанные в этот период модели показали, что в случайным образом генерируемой сети взаимодействия в сообществе при удалении бессмысленных цепей (типа А ест В, В ест С, С ест А и подобного типа) локальная устойчивость падает с увеличением сложности. Если продолжить усложнение модели и учесть, что консументы испытывают влияние пищевых ресурсов, а пищевые ресурсы от консументов не зависят, то можно прийти к выводу о том, что устойчивость не зависит от сложности, либо также падает с её увеличением. Разумеется, такие результаты справедливы главным образом для детритных цепей питания, в которых консументы не влияют на поток пищевых ресурсов , хотя и могут менять пищевую ценность последних.
При изучении общей устойчивости на модели из 6 видов (2 хищника-консумента второго порядка, 2 консумента первого порядка и 2 вида в основании пищевой цепи) исследовалось удаление одного из видов. В качестве параметра устойчивости была принята связность. Сообщество считалось устойчивым, если остальные виды оставались локально устойчивыми. Полученные результаты согласовывались с общепринятыми воззрениями о том, что с повышением сложности при выпадении хищников высшего порядка устойчивость сообщества падает, но при выпадении оснований пищевой цепи с повышением сложности устойчивость повышалась .
В случае , когда под сложностью также понимается связность, с повышением сложности упругая устойчивость также повышается. То есть, большее разнообразие видов и большая сила связи между ними позволяет сообществам быстрее восстанавливать свою структуру и функции. Данный факт подтверждает общепринятые взгляды на роль биоразнообразия как некоего пула (фонда) для восстановления полноценной структуры как экосистем, так и более высокоорганизованных структур биосферы, а также самой биосферы в целом. На данный момент общепринятым и фактически неоспариваемым является представление о том, что биосфера эволюционировала в сторону увеличения биоразнообразия (всех трёх его компонентов), ускорения обращения вещества между компонентами биосферы, и «убыстрения» времени жизни как видов, так и экосистем .

На данный момент научное понимание всех процессов внутри экосистемы далеко от совершенства, и в большей части исследований либо вся экосистема, либо некоторые её части выступают в качестве « чёрного ящика » . В то же время, как любая относительно замкнутая система, экосистема характеризуется входящим и выходящим потоком энергии и распределением этих потоков между компонентами экосистем.
При анализе продуктивности и потоков вещества и энергии в экосистемах выделяют понятия биомасса и урожай на корню . Под урожаем на корню понимается масса тел всех организмов на единице площади суши или воды , а под биомассой — масса этих же организмов в пересчёте на энергию (например, в джоулях ) или в пересчёте на сухое органическое вещество (например, в тоннах на гектар ) . К биомассе относят тела организмов целиком, включая и витализированные омертвевшие части и не только у растений, к примеру, кора и ксилема , но и ногти и ороговевшие части у животных. Биомасса превращается в некромассу только тогда, когда отмирает часть организма (отделяется от него) или весь организм. Часто зафиксированные в биомассе вещества являются «мёртвым капиталом», особенно это выражено у растений: вещества ксилемы могут сотнями лет не поступать в круговорот, служа только опорой растения .
Под первичной продукцией сообщества (или первичной биологической продукцией) понимается образование биомассы (более точно — синтез пластических веществ) продуцентами без исключения энергии, затраченной на дыхание за единицу времени на единицу площади (например, в сутки на гектар).
Первичную продукцию сообщества разделяют на валовую первичную продукцию , то есть всю продукцию фотосинтеза без затрат на дыхание , и чистую первичную продукцию , являющуюся разницей между валовой первичной продукцией и затратами на дыхание. Иногда её ещё называют чистой ассимиляцией или наблюдаемым фотосинтезом ).
Чистая продуктивность сообщества — скорость накопления органического вещества, не потребляемого гетеротрофами (а затем и редуцентами). Обычно вычисляется за вегетационный период либо за год . Таким образом, это часть продукции, которая не может быть переработана самой экосистемой. В более зрелых экосистемах значение чистой продуктивности сообщества стремится к нулю (см. концепцию ).
Вторичная продуктивность сообщества — скорость накопления энергии на уровне консументов. Вторичную продукцию не подразделяют на валовую и чистую, так как консументы только потребляют энергию, усвоенную продуцентами, часть её не ассимилируется, часть идёт на дыхание, а остаток идёт в биомассу, поэтому более корректно называть её вторичной ассимиляцией .

Распределение энергии и вещества в экосистеме может быть представлено в виде системы уравнений. Если продукцию продуцентов представить как P 1 , то продукция консументов первого порядка будет выглядеть следующим образом:
где R 2 — затраты на дыхание, теплоотдача и неассимилированная энергия. Следующие консументы (второго порядка) переработают биомассу консументов первого порядка в соответствии с:
и так далее, до консументов самого высшего порядка и редуцентов. Таким образом, чем больше в экосистеме потребителей (консументов), тем более полно перерабатывается энергия, первоначально зафиксированная продуцентами в пластических веществах . В климаксных сообществах, где разнообразие для данного региона обычно максимально, такая схема переработки энергии позволяет сообществам устойчиво функционировать на протяжении длительного времени.

Хозяйственная деятельность человека изменяет величину чистой первичной продукции экосистем. В некоторых регионах, примером которых может служить долина Нила , ирригация дает значительное увеличение локальной чистой первичной продукции, но это скорее исключение из правила. Глобально на поверхности суши антропогенное воздействие сокращает этот показатель на 9,6 % . 23,8 % чистой первичной продукции биосферы человечество потребляет на свои нужды . Это является следствием того, что 34 % (2000 год) всей свободной ото льда поверхности суши заняты под нужды сельского хозяйства , в том числе 12 % под пашни и 22 % под пастбища . Это сокращает количество энергии, доступной для других биологических видов, негативно влияет на биоразнообразие, затрагивает водный и углеродный циклы планеты. Дальнейшее увеличение этой доли создает угрозу разрушения жизненно важных экосистем планеты .

В природе, как правило, не существует чётких границ между различными экосистемами . Всегда можно указать на ту или иную экосистему, но выделить дискретные границы, если они не представлены различными ландшафтными факторами (обрывы, реки, различные склоны холмов, выходы скальных пород и т. п.), не представляется возможным, ибо чаще всего существуют плавные переходы от одной экосистемы к другой . Это обусловлено относительно плавным изменением градиента факторов среды (влажность, температура, увлажнённость и прочее). Иногда переходы из одной экосистемы в другую могут фактически являться самостоятельной экосистемой. Обычно сообщества, образующиеся на стыке различных экосистем, называются экотонами . Термин «экотон» введён Ф. Клементсом в 1905 году.
Экотоны играют существенную роль в поддержании биологического разнообразия экосистем за счёт так называемого краевого эффекта — сочетания комплекса факторов среды различных экосистем, обуславливающее большее разнообразие условий среды, следовательно, и экологических ниш . Тем самым возможно существование видов как из одной, так и из другой экосистемы, а также специфичных для экотона видов (например ).

В российской литературе краевой эффект иногда называют эффектом опушки .
Примерами экотонов могут служить прибрежные участки суши и водоёмов (например, литораль ), опушки, переходы из лесных экосистем в полевые, эстуарии . Однако не всегда экотон является местом повышенного биоразнообразия видов. К примеру, эстуарии рек, впадающих в моря и океаны , наоборот, характеризуются пониженным биоразнообразием видов, так как средняя солёность дельт не позволяет существовать многим пресноводным и солоноводным (морским) видам.
Альтернативным представлением о континуальных переходах между экосистемами является представление о экоклинах (экологических рядах). Экоклин — постепенная смена биотопов, генетически и фенотипически приспособленных к конкретной среде обитания, при пространственном изменении какого-либо фактора среды (обычно климатического), а потому составляющих непрерывный ряд форм без заметных перерывов постепенности. Экоклин невозможно разделить на экотипы. Например, длина ушей лисиц и мн. др., их признаки изменяются с севера на юг настолько постепенно, что очень затруднительно выделить четкие морфологические группы, которые бы естественно объединялись в подвиды.

На одном и том же биотопе с течением времени существуют различные экосистемы. Смена одной экосистемы на другую может занимать как довольно длительные, так относительно короткие (несколько лет) промежутки времени. Длительность существования экосистем в таком случае определяется этапом сукцессии . Смена экосистем в биотопе может быть обусловлена и катастрофическими процессами, но в таком случае, существенно изменяется и сам биотоп, и такую смену не принято называть сукцессией (за некоторыми исключениями, когда катастрофа, например, пожар — естественный этап циклической сукцессии) .
Сукцессия — это последовательная, закономерная смена одних сообществ другими на определённом участке территории, обусловленная внутренними факторами развития экосистем . Каждое предыдущее сообщество предопределяет условия существования следующего и собственного исчезновения . Это связано с тем, что в экосистемах, которые являются переходными в , происходит накопление вещества и энергии, которые они уже не в состоянии включить в круговорот, преобразование биотопа, изменение микроклимата и других факторов, и тем самым создаётся вещественно-энергетическая база, а также и условия среды, необходимые для формирования последующих сообществ. Однако, есть и другая модель, которая объясняет механизм сукцессии следующим образом : виды каждого предыдущего сообщества вытесняются лишь последовательной конкуренцией, ингибируя и «сопротивляясь» внедрению последующих видов. Тем не менее, эта теория рассматривает лишь конкурентные отношения между видами, не описывая всю картину экосистемы в целом. Безусловно, такие процессы идут, но конкурентное вытеснение предыдущих видов возможно именно из-за преобразования ими биотопа. Таким образом, обе модели описывают разные аспекты процесса и верны одновременно.

Сукцессия бывает автотрофной (например, сукцессия после лесного пожара) и гетеротрофной (например, осушенное болото ) . На ранних стадиях автотрофной сукцессионной последовательности соотношение P/R много больше единицы, так как обычно первичные сообщества обладают высокой продуктивностью, но структура экосистемы ещё не сформировалась полностью, и нет возможности утилизировать эту биомассу . Последовательно, с усложнением сообществ, с усложнением структуры экосистемы, расходы на дыхание (R) растут, так как появляется всё больше гетеротрофов, ответственных за перераспределение вещественно-энергетических потоков, соотношение P/R стремится к единице и фактически является таковым у терминального сообщества (экосистемы) . Гетеротрофная сукцессия обладает обратными характеристиками: в ней соотношение P/R на ранних этапах много меньше единицы (так как существует много органического вещества и нет необходимости в его синтезе, его можно сразу использовать на построение сообщества) и постепенно увеличивается по мере продвижения по сукцессионным стадиям.

На ранних этапах сукцессии видовое разнообразие мало, но по мере развития разнообразие нарастает, изменяется видовой состав сообщества, начинают преобладать виды со сложными и продолжительными жизненными циклами, обычно появляются всё более крупные организмы, происходит развитие взаимовыгодных коопераций и симбиозов , усложняется трофическая структура экосистемы. Обычно предполагается, что терминальная стадия сукцессии обладает наибольшим видовым биоразнообразием. Это справедливо не всегда, но для климаксных сообществ тропических лесов это утверждение справедливо , а для сообществ умеренных широт пик разнообразия приходится на середину сукцессинного ряда или ближе к терминальной стадии . На ранних стадиях сообщества состоят из видов с относительно высокой скоростью размножения и роста, но низкой способностью к индивидуальному выживанию (r-стратеги). В терминальной стадии воздействие естественного отбора благоприятствует видам с низкой скоростью роста, но большей способностью к выживанию (k-стратеги).
По мере продвижения по сукцессионному ряду происходит всё большее вовлечение биогенных элементов в круговорот в экосистемах, возможно относительное замыкание внутри экосистемы потоков таких биогенных элементов, как азот и кальций (одни из наиболее подвижных биогенов ) . Поэтому в терминальной стадии, когда большая часть биогенов вовлечена в круговорот, экосистемы более независимы от внешнего поступления данных элементов .
Для исследования процесса сукцессии применяют различные математические модели, в том числе

Понятие сукцессии тесно связано с понятием климаксного сообщества . Климаксное сообщество формируется в результате последовательной смены экосистем и представляет собой наиболее сбалансированное сообщество , максимально эффективно использующее вещественно-энергетические потоки, то есть поддерживающее максимально возможную биомассу на единицу поступающей в экосистему энергии.

Теоретически у каждого сукцессионного ряда существует климаксное сообщество (экосистема), которое является терминальной стадией развития (или несколько, так называемая концепция поликлимакса). Однако, в реальности сукцессинный ряд замыкается климаксом не всегда, может реализоваться субклимаксное сообщество (или названное Ф. Клементсом — плагиклимакс), которое представляет собой сообщество, предшествующее климаксному, достаточно развитое структурно и функционально . Такая ситуация может возникать в силу естественных причин — условий среды или вследствие деятельности человека (в таком случае его называют дисклимакс ).
Вопрос ранжирования экосистем достаточно сложен. Выделение минимальных экосистем (биогеоценозов) и экосистемы наивысшего ранга — биосферы не вызывает сомнений . Промежуточные же выделения довольно сложны, так как сложности не всегда однозначно позволяют определить границы экосистем. В геоэкологии (и ландшафтоведении ) существует следующее ранжирование: фация — урочище (экосистема) — ландшафт — — географическая область — биом — биосфера . В экологии существует сходное ранжирование , однако, обычно считается, что корректно выделение лишь одной промежуточной экосистемы — биома.
Биом — крупное системно-географическое (экосистемное) подразделение в пределах природно-климатической зоны ( Реймерс Н. Ф. ). Согласно Р. Х. Уиттекеру — группа экосистем данного континента, которые имеют сходную структуру или физиономию растительности и общий характер условий среды. Это определение несколько некорректно, так как существует привязка к конкретному континенту, а некоторые биомы присутствуют на разных континентах, например, тундровый биом или степной .
На данный момент наиболее общепринятое определение звучит так: «Биом — совокупность экосистем со сходным типом растительности, расположенных в одной природно-климатической зоне» ( , ).
Общим в этих определениях является то, что в любом случае биомом называется совокупность экосистем одной природно-климатической зоны.
Выделяют от 8 до 30 биомов. Географическое распределение биомов определяют :
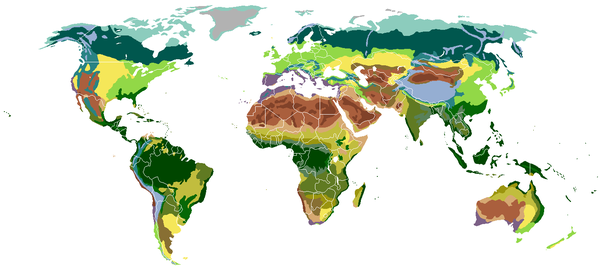
| Наземные биомы, классифицированные по типу растительности | ||||
| Полярные пустыни Тундра Тайга Широколиственные леса | Степи Субтропические дождевые леса Средиземноморские биомы Муссонные леса | Аридные пустыни Ксерофитных кустарников Южные степи Семиаридные пустыни | Саванны Саванны с древесной растительностью (лесостепи) Субтропический лес Тропический дождевой лес | Альпийская тундра Горные леса |

Термин биосфера был введён Жаном-Батистом Ламарком в начале XIX века, а в геологии предложен австрийским геологом Эдуардом Зюссом в 1875 году . Однако создание целостного учения о биосфере принадлежит русскому учёному Владимиру Ивановичу Вернадскому .
Биосфера — экосистема высшего порядка, объединяющая все остальные экосистемы и обеспечивающая существование жизни на Земле. В состав биосферы входят следующие «сферы» :
Биосфера тоже не замкнутая система, она фактически полностью обеспечивается энергией Солнца , небольшую часть составляет тепло самой Земли. Ежегодно Земля получает от Солнца около 1,3⋅10 24 калорий . 40 % от этой энергии излучается обратно в космос, около 15 % идёт на нагрев атмосферы, почвы и воды, вся остальная энергия является видимым светом, который и является источником фотосинтеза.
В. И. Вернадский впервые чётко сформулировал понимание того, что всё живое на планете неразрывно связано с биосферой и обязано ей своим существованием:
В действительности, ни один живой организм в свободном состоянии на Земле не находится. Все эти организмы неразрывно и непрерывно связаны — прежде всего питанием и дыханием — с окружающей их материально-энергетической средой. Вне её в природных условиях они существовать не могут.
— В. И. Вернадский
Живое вещество (совокупность всех организмов на Земле) составляет ничтожно малую часть от массы Земли, однако влияние живого вещества на процессы преобразования Земли огромно . Весь тот облик Земли, который наблюдается сейчас, не был бы возможен без миллиардов лет жизнедеятельности живого вещества.
На данный момент сам человек, как часть живого вещества, является существенной геологической силой и значительно изменяет направления процессов, происходящих в биосфере, тем самым ставя под угрозу своё существование:
В ярком образе экономист Л. Брентано иллюстрировал планетную значимость этого явления. Он подсчитал, что, если бы каждому человеку дать один квадратный метр и поставить всех людей рядом, они не заняли бы даже всей площади маленького Боденского озера на границе Баварии и Швейцарии. Остальная поверхность Земли осталась бы пустой от человека. Таким образом, всё человечество, вместе взятое, представляет ничтожную массу вещества планеты. Мощь его связана не с его материей, но с его мозгом, с его разумом и направленным этим разумом его трудом.
…
В гуще, в интенсивности и в сложности современной жизни человек практически забывает, что он сам и всё человечество, от которого он не может быть отделён, неразрывно связаны с биосферой — с определённой частью планеты, на которой они живут. Они — геологически закономерно связаны с её материально-энергетической структурой.
…
Человечество, как живое вещество, неразрывно связано с материально-энергетическими процессами определённой геологической оболочки Земли — с её биосферой. Оно не может физически быть от неё независимым ни на одну минуту.
…
Лик планеты — биосфера — химически резко меняется человеком сознательно и главным образом бессознательно . Меняется человеком физически и химически воздушная оболочка суши, все её природные воды.
— В. И. Вернадский .

Искусственные экосистемы — это экосистемы, созданные человеком, например, агроценозы , природно-хозяйственные системы или Биосфера 2 .
Искусственные экосистемы имеют тот же набор компонентов, что и естественные: продуценты , консументы и редуценты , но есть существенные отличия в перераспределении потоков вещества и энергии. В частности, созданные человеком экосистемы отличаются от естественных следующим :
Без поддержания энергетических потоков со стороны человека в искусственных системах с той или иной скоростью восстанавливаются естественные процессы и формируется естественная структура компонентов экосистемы и вещественно-энергетических потоков между ними.
В данных науках существуют понятия, схожие с понятием экосистема. Различие состоит в том, что в данных науках происходит смещение аспекта рассмотрения структуры и функций экосистем.
В экогеологии , науке, изучающей взаимодействие организмов и литосферы, упор делается на рассмотрении данных связей и соответственно взаимодействии данных компонентов природы, причём в большей степени влияние литосферы на формирование, развитие и функционирование экосистем и отдельных компонентов.
Для ландшафтоведения экосистема представляет интерес как системная единица, занимающая некоторое урочище и формирующая уникальный «рисунок» ландшафта .
В геоэкологии основное внимание уделяется изучению связей между равноправными компонентами геосистемы , так как геосистема полицентрична, в то время как экосистема моноцентрична и основной компонент в ней — биота .
В целом, в географических науках принято рассматривать природный территориальный комплекс , как эквивалент экосистемы.
|
|
|
|---|---|
| Природные зоны | |
| Функциональные компоненты | |
| Структурные компоненты | |
| Абиотические компоненты | |
| Функционирование | |
| Загрязнение экосистем | |
| История Земли |
|
|
|---|---|---|
| Физические свойства Земли | ||
| Оболочки Земли | ||
|
География
и геология |
||
| Окружающая среда | ||
| См. также | ||
|
||
|
|
Эта статья входит в число
хороших статей
русскоязычного раздела Википедии.
|
|
|
Некоторые
внешние ссылки
в этой статье
ведут на сайты, занесённые в
спам-лист
|
| Экосистема | |
|---|---|

|
|
| Изучается в | экология , науки о жизни и |
| Предсказано кем | Артур Рой Клэпем |




