Interested Article - Бескишечные турбеллярии
- 2020-04-22
- 1
Бескише́чные турбелля́рии ( лат. Acoela ) — класс животных из подтипа Acoelomorpha типа Xenacoelomorpha (некоторые систематики понижают ранг таксона Acoela до отряда ), включает около 400 видов . Ранее рассматривались как отряд ресничных червей (Turbellaria) . Это небольшие (до 9 мм в длину) животные, не имеющие ни целома , ни кишки. Строение бескишечных турбеллярий включает в себя покрытый ресничками и богатый железами эпидермис , кольцевую и продольную мускулатуру , нервную систему с примитивным мозгом и паренхиму , в которую заключены внутренние органы животного . Роль пищеварительной системы у этих животных выполняет центральный синцитий , в котором пища усваивается посредством фагоцитоза . Непереваренные остатки пищи выводятся наружу через ротовое отверстие . У бескишечных турбеллярий есть органы чувств, такие как статоцист и фоторецепторы . У этих животных имеется весьма сложная (как правило, гермафродитная ) репродуктивная система . Acoela могут размножаться как бесполым путём , так и половым , причём в последнем случае оплодотворение всегда внутреннее, а развитие потомства прямое (в редких случаях имеет место живорождение). Кроме того, животные из этой группы обладают способностью регенерировать , создавая две полноценные особи из одной разрезанной .
Бескишечные турбеллярии обитают во всех океанах, большинство видов — свободноживущие обитатели бентоса . Некоторые виды обитают на кораллах или являются комменсалами других беспозвоночных, а некоторые имеют собственных эндосимбионтов .
Систематика бескишечных турбеллярий и их местоположение в царстве животных несколько раз пересматривались. Исследования генома разных видов Acoela методами молекулярного анализа указывают на то, что этот класс относится к кладе , лежащей в основании всех двусторонне-симметричных животных .
Описание
Внешний вид

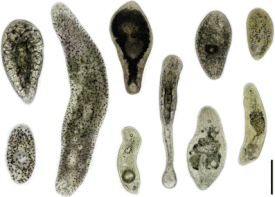
Acoela — мелкие многоклеточные животные, двусторонне-симметричные черви, как правило, их размер не превышает 2 мм , но крупные представители некоторых видов из семейства Convolutidae , могут достигать и 9 мм в длину . Форма тела зависит от местообитания: у обитателей песка тело длинное и тонкое, живущие в иле имеют компактное тело в форме капли, живущие между камнями и кораллами широкие и плоские, у эпифитных видов края тела подвёрнуты к брюшной стороне, пелагические виды имеют дисковидное тело, часто с подвёрнутыми краями. Большинство видов Acoela — это полупрозрачные животные, но встречаются виды, представители которых окрашены в различные цвета, например зелёные Symsagittifera roscoffensis (окраска вызвана симбионтом ) или красные (из-за наполненных гемоглобином пигментных клеток) . У турбеллярий нет специальных локомоторных придатков. Как и настоящие мелкие турбеллярии, бескишечные турбеллярии перемещаются, скользя по субстрату или плавая в воде с помощью биения ресничек . Ресничная локомоция может быть весьма эффективной, позволяя, в том числе, резкое изменение движения на 180 градусов .
Эпидермис

Внешняя поверхность бескишечных турбеллярий представляет собой однослойный ресничный эпидермис . У многих видов Acoela клетки эпидермиса оказываются разделёнными мышечным поясом на две части: внешнюю, широкую и плоскую, и внутреннюю, в которой находится ядро клетки. Однако у видов и такое строение эпидермальных клеток не было показано, у этих видов я́дра клеток эпидермиса находятся над поясом мышц. Каждая клетка эпидермиса несёт большое количество ресничек . В терминальных частях эпителиальные клетки соединены десмосомами . Между ресничками могут быть микроворсинки . Реснички бескишечных турбеллярий, как и у других Acoelomorpha, отличаются особым ступенчатым строением апикальной области. Корешки ресничек также представляют собой сложную структуру, состоящую из двучленного основного корешка и двух дополнительных корешков . Таким образом, корешки создают разветвлённую систему из микротрубочек , которая соединяет соседние реснички . У этих животных эпидермис не покрыт кутикулой. Базальная мембрана и внеклеточный матрикс под эпидермисом у Acoela практически отсутствуют, и прямо под эпидермисом располагается слой кольцевой мускулатуры . Эпидермис может нести дополнительные структуры. Например, у вида Paratomella rubra на заднем конце тела имеются специальные реснички, позволяющие представителям этого вида временное прикрепление к субстрату. В отличие от обычных ресничек, свободные концы прикрепительных ресничек уплощены . Эпидермис бескишечных турбеллярий не несёт стволовых клеток . Обновление клеток эпидермиса происходит за счёт пролиферации стволовых клеток, расположенных в паренхиме, прямо под эпидермисом .
В эпидермисе находятся многочисленные одноклеточные железы. Среди них рабдоидные железы, которые могут быть окрашены, и слизистые железы. На апикальном полюсе тела червя расположено скопление из двух или нескольких крупных желёз, секретирующих слизь и открывающихся фронтальной по́рой. Это скопление получило название фронтального органа . Рядом с фронтальным органом залегает скопление слизистых желёз, которые открываются единым отверстием на переднем конце тела. На нём же располагаются разнообразные фронтальные железы. Ядра железистых клеток, за исключением большинства пигментных клеток, находятся под слоем мускулатуры . Бескишечные турбеллярии семейства Sagittiferidae несут на поверхности тела особые уникальные игольчатые структуры — сагиттоцисты. Их выделяют специализированные железистые клетки — сагиттоциты благодаря сокращению специальных мышц. Сагиттоциты могут быть связаны с половым поведением, а также служить для защиты и охоты .
Паренхима

Паренхима бескишечных турбеллярий является соединительной тканью между эпидермальным слоем и пищеварительным синцитием. В отличие от представителей других близкородственных таксонов, у которых клетки паренхимы заключены в слой внеклеточного матрикса , у Acoela паренхима практически целиком состоит из клеток при минимальном количестве внеклеточного матрикса . Последние исследования показали, что паренхима Acoela и паренхима плоских червей не гомологичны друг другу и являются результатом параллельной эволюции . В паренхиме можно выделить периферическую и центральную зоны (в состав последней также входит пищеварительный синцитий ). Паренхима состоит из нескольких типов клеток с разными функциями. Один из типов клеток — это уже упомянутые выше стволовые клетки (необласты), которые при необходимости дифференциируются и мигрируют в область эпидермиса. Каждая такая клетка несёт в себе пучок центриолей , из которых в процессе дифференциации образуются новые эпидермальные реснички. Стволовые клетки Acoela способны к перемещениям. Необласты Acoela служат единственным источником как для соматических клеток , так и для клеток зародышевой линии . Другой тип клеток паренхимы — стационарные клетки. Эти клетки обладают разветвлённым телом, что позволяет им создавать щелевые контакты как между собой, так и с клетками всех прочих тканей животного. Щелевые контакты обеспечивают быстрый транспорт веществ внутри Acoela и служат альтернативой отсутствующей у этой группы животных системы циркуляции. Кроме этих двух типов паренхимных клеток, наличествующих у всех бескишечных турбеллярий, у некоторых видов можно обнаружить дополнительные типы клеток. Например, у вида Paratomella rubra , в полном соответствии с видовым эпитетом , тело окрашено в красный цвет за счёт пигментных паренхимных клеток, содержащих гемоглобин . Иногда клетки паренхимы содержат вакуоли .
Мускулатура

У Acoela имеются только гладкие мышцы . Однако у некоторых видов в миофибриллах наблюдается поперечная исчерченность, похожая на исчерченность и Z-диски настоящих поперечнополосатых мышц . Длина « саркомера » в этом случае составляет около 1,5 мкм. Любопытно, что распределение таких мышечных белков , как актин , и тропомиозин в мышцах этих червей соответствует поперечнополосатой мускулатуре остальных Bilateria .
У большинства видов бескишечных турбеллярий внешние мышцы располагаются по следующей схеме. Прямо под стенкой тела (эпидермисом) залегают кольцевые мышцы, под ними — диагональные, а слоем ниже продольные мышцы. Единственным исключением является семейство , у представителей которого внешний (близкий к эпидермису) слой мышц продольный, а под ним располагается кольцевой. Диагональные мышцы пересекаются сразу за ротовым отверстием , обвиваясь вокруг него и создавая на пересечении U-образную мышцу (за исключением семейства , у которого эта структура отсутствует). Количество и расположение дорсальных и вентральных диагональных мышц варьирует от семейства к семейству. У некоторых видов Acoela, например, Symsagittifera roscoffensis и вокруг рта дополнительно образуется сфинктер . В отличие от достаточно постоянной схемы расположения внешних мышц, разные виды Acoela демонстрируют весьма различный с точки зрения разветвлённости и расположения набор внутренних мышц (мышц паренхимы ). Основные внутренние мышцы бескишечных турбеллярий делятся на мышцы статоциста (которые удерживают его на месте), дорсовентральные мышцы и мышцы копулятивных органов . Наиболее развитая мышечная система была описана для более крупных видов Acoela, таких как Symsagittifera roscoffensis , а наименее развитая у представителей рода Childia ( ) и у, предположительно, наиболее базального таксона класса — .
Нервная система
Бескишечные турбеллярии обладают примитивным мозгом, который имеет форму кольца и расположен под эпидермисом в передней части животного. Мозг не отделён от остального тела мембраной, и сквозь него проходят мышечные пучки и протоки фронтальных желёз. От мозга отходят к задней части тела несколько нервных тяжей, расположенных радиально-симметрично. Нервные тяжи соединены с периферийной нервной системой, состоящей из трёх обособленных нервных сетей, которые находятся под слоем мускулатуры (расположенной непосредственно под стенкой тела), между слоем мышц и эпидермисом и в самом эпидермисе соответственно. Такое распределение нервной системы обеспечивает хорошую иннервацию всех слоёв мускулатуры . Нервная система лучше развита на спинной стороне тела червя, чем на брюшной. С помощью методов иммуногистохимии в нервной системе Acoela были найдены серотониноподобные иммунореактивные, и холинергические части .
Органы чувств

Все известные виды Acoela обладают одним статоцистом , который служит рецептором силы тяжести . Статоцист расположен в передней части тела и состоит из литоцита, имеющего один статолит и заключённого в капсулу из двух париетальных клеток . Предположительно, основной функцией статоциста является регулирование равновесия животного. Статоцист служит отличительной чертой таксона. Иногда особи, появившиеся на свет с помощью бесполого размножения , не имеют статоциста, а у некоторых червей после регенерации переднего конца тела имеется несколько статоцистов .
У некоторых видов на переднем конце тела имеются парные глазные пятна, по-видимому, служащие фоторецепторами . Так, у глазные пятна представляют собой специализированные эпидермальные клетки с ресничками обычного строения и многочисленными пигментными гранулами. На базальной стороне этих клеток имеется множество синапсов , однако фоторецепторная функция этих образований не доказана. Она показана для глазных пятен . У этого и некоторых других видов в глазных пятнах нет ресничек; они состоят из пигментных клеток, содержащих вакуоли со светопреломляющими включениями, и до трёх нервных клеток , проводящих сигнал. Многие Acoela могут чувствовать свет при помощи фоторецепторных клеток эпидермиса, которые очень трудно идентифицировать, так как они не окружены пигментными клетками . Проведённые на бескишечных турбелляриях эксперименты продемонстрировали отрицательный фототаксис для представителей этого класса (в описанных экспериментах животные стремились удалиться от источника света), что позволяет предположить, что для них свет сигнализирует угрозу .
Рядом со ртом и в глотке (если она есть) находятся одножгутиковые рецепторные клетки . Описано несколько морфологических типов этих рецепторов, и разные семейства имеют разные типы рецепторов .
Пищеварительная система
Пищеварительная система бескишечных тубеллярий состоит из ротового отверстия и центрального пищеварительного синцития , который у некоторых представителей связан с мышцами . Полости кишки у этих животных нет, за что они и получили своё название . У одной из наиболее примитивных бескишечных турбеллярий, Paratomella rubra , нет и синцития, и паренхима состоит только из отдельных клеток . Поглощаемая пища переходит из ротовой полости в центральный синцитий путём фагоцитоза , отходы пищеварения выделяются через ротовое отверстие. Таким образом, пищеварение у этих животных не полостное, а внутриклеточное. Пищеварительные железы на синцитии отсутствуют. Как правило, у представителей этого класса животных нет ни клеточной кишки , ни ануса . Наличие глотки показано для семейств Diopisthoporidae , , , Solemofilomorphidae, а также для родов и . Интересно, что мускулистые глотки у разных таксонов Acoela не гомологичны друг другу и сильно различаются анатомически . Более того, они не гомологичны и глотке плоских червей . У некоторых Acoela, например, и ряда представителей семейства , имеется единственная пищеварительная полость, не выстланная эпителием. Она окружена временными синцитиальными структурами и наполнена окружёнными мембраной везикулами; кроме того, она связана с мышечными образованиями .
Расположение рта варьируется у разных видов. Он неизменно находится на средней линии тела, на вентральной поверхности, но на этой линии может находиться как у переднего края животного (например, у ), так и у заднего (например, у Diopisthoporus ) или где-либо посередине .
Выделительная система
У Acoela не было найдено типичных органов выделения . Были найдены клетки, напоминающие циртоциты протонефридиев , однако впоследствии оказалось, что они представляют собой дегенерирующие эпидермальные клетки. Клетки, лишённые ресничек и похожие на канальцевые клетки протонефридиев, с разветвлёнными лакунами и трубочками, были обнаружены у Paratomella rubra . Возможно, они выполняют выделительную функцию .
Внутренний транспорт
Обмен газов бескишечные турбеллярии осуществляют всей поверхностью тела. Их уплощённое строение в дорсо-вентральном направлении позволяет осуществлять эффективную доставку кислорода во внутренние ткани и вывод из них углекислого газа посредством диффузии . Доставка питательных веществ из центрального синцития во внешние ткани может также осуществляться за счёт внутриклеточной диффузии или альтернативно транспортироваться через межклеточные щелевые контакты .
Репродуктивная система

Репродуктивная система бескишечных турбеллярий значительно отличается от семейства к семейству и до применения молекулярных методов кладистики служила основным морфологическим признаком, по которому производилась классификация представителей этого класса. Практически все виды бескишечных турбеллярий — гермафродиты , нередко протандрические, то есть особь сначала функционирует как самец, а затем — как самка .
Мужская половая система обеспечивает продукцию сперматозоидов , агрегацию их в сперме и выведение спермы из организма. В неё могут входить парные семенники , семяпроводы, семенной пузырёк, в котором запасается собственная сперма животного, , в которой образуется секрет, и орган совокупления — пенис . Строение пениса очень разнообразно: у некоторых видов его нет, у других копулятивный орган представляет собой простую инвагинацию стенки тела, а у некоторых пенис имеет очень сложное строение и включает мышечные, склеротизированные и железистые элементы. У некоторых видов пенис несёт на себе стилет, с помощью которого в ходе гиподермального оплодотворения прокалывается эпидермис партнёра . Сперма обычно образует скопление в паренхиме рядом с копулятивным органом. Иногда эти скопления окружаются специализированной тканью, содержащей мышечные элементы, которые способствуют выпрыскиванию спермы; в таком случае скопления спермы называют семенными везикулами .
Женская половая система обеспечивает продуцирование яиц, их развитие, хранение и вывод наружу после оплодотворения, а также поступление полученной от партнёра спермы внутрь. У некоторых видов Acoela женская половая система не имеет выводного отверстия. У других же она открывается наружу женским половым отверстием, так называемым гонопором (в некоторых источниках используется термин « » или «влагалище»). Поступающая от партнёра сперма может недолго храниться в копулятивной сумке (иногда используется термин « » или «женский копуляторный орган» ), а если необходимо долгосрочное хранение, сперма перенаправляется в . Нередко имеются один или несколько бурсальных носиков ( англ. nozzles ). Эти склеротизированные структуры укреплены клетками, богатыми фибриллярным актином , и тесно связаны с копулятивной бурсой, принимая участие в модификации спермы. Детали их строения использовались для разработки классификации Acoela . У Acoela нет яйцеводов, для вывода яиц у некоторых видов используется ротовая полость, а у других образуются временные разрывы стенки тела . Ооциты нередко окружены дополнительными клетками .
Гонады бескишечных турбеллярий не имеют эпителия. Клетки зародышевой линии развиваются в паренхиме и никак не отделяются от неё. Расположение гонад в теле может сильно разниться даже у особей одного вида; яичники и семенники могут быть парными или непарными, а у некоторых червей, например, Antigonaria , нет отдельных гонад , вместо них есть герминативная зона, продуцирующая как сперматозоиды, так и яйцеклетки .
Строение сперматозоидов имеет важное значение для определения внутренней филогении Acoela . Сперматозоиды этих животных не имеют свободных жгутиков , а только лишь две аксонемы , полностью погружённые в тело сперматозоида . Аксонемы, как правило, имеют типичное количество микротрубочек : 9 периферических пар микротрубочек и две центральные микротрубочки; впрочем, иногда имеется только одна центральная микротрубочка или центральные микротрубочки полностью отсутствуют. В цитоплазме сперматозоида имеются микротрубочки, придающие упругость клетке . Сперматозоиды червей из разных семейств могут также различаться отношением длины ядра к длине всей клетки, положением ядра и перекрыванием ядра и жгутика .
Симбионты

У многих Acoela из семейств Convolutidae и Sagittiferidae , населяющих местообитания, освещаемые солнцем, имеются симбиотические водоросли , необходимые для выживания хозяина. Это могут быть зоохлореллы или . Например, в клетках паренхимы червя Symsagittifera roscoffensis живут зелёные водоросли Tetraselmis convolutae , которые придают телу червя зелёный цвет . Симбионты передаются преимущественно горизонтально; это означает, что каждое новое поколение приобретает симбионтов заново. В этом случае черви рождаются бесцветными и в течение нескольких дней после рождения заглатывают свободноживущие клетки водорослей, снабжённые жгутиками. Особи, не сумевшие поймать водоросль, умирают. После поглощения клетки водорослей попадают под эпидермис и проникают внутрь клеток червя, меняя форму и утрачивая жгутик, теку и глазок . У Amphiscolops carvalhoi и Waminoa brickneri описана вертикальная передача симбионтов, при которой клетки водорослей содержатся в яйце и передаются следующему поколению. Симбиоз с водорослями у Acoela появлялся минимум дважды . В некоторых случаях эндосимбионты полностью обеспечивают своего хозяина, и сами черви не питаются. Черви Convolutriloba longifissura не выживают без симбионтов, однако продолжают также питаться самостоятельно .
Размножение
Половое размножение
Как упоминалось выше, бескишечные турбеллярии, как правило, являются гермафродитами. Тем не менее, самооплодотворение одной особи исключено в силу строения половой системы. Оплодотворение у Acoela всегда внутреннее. Способ копуляции может отличаться от вида к виду и значительно зависит от окружающей среды. Возможен взаимный обмен спермой, гипердермальная передача и гиподермальное введение. В первых двух случаях в передаче спермы участвуют простое отверстие в эпидермисе — антрум или мягкий, мускулистый пенис . В последнем случае эпидермис партнёра прокалывается склеротизированными вспомогательными структурами, похожими на иголки или стилеты . Количество яиц, производимое каждой особью, небольшое, так как минимальный размер яйца составляет около 50 мкм, что для животных с размерами в несколько миллиметров является существенным ограничивающим фактором . Яйца откладываются поодиночке или группами, окружёнными слизью , через рот, женский гонопор или посредством разрушения клеточной стенки с последующим прямым развитием особи . Некоторые представители, например, Diopisthoporus и Childia vivipara — живородящие .
Бесполое размножение

В то время как половое размножение известно у всех Acoela, многие из них также способны к бесполому размножению . У семейства Paratomellidae описана — образование цепочки зооидов, которая потом разрывается с образованием дочерних особей. В пределах семейства Convolutidae распространена архитомия , или фрагментация, при которой недостающие органы формируются после разделения материнской и дочерней особей. У происходит почкование, при котором дочерняя особь развивается из одной из трёх лопастей, расположенных на заднем конце материнской особи, в направлении, перпендикулярном или обратном материнской особи .
Регенерация
У Acoela ярко выражена способность к регенерации после разделения одной особи на две. У всех видов , изученных на данный момент, в начале процесса происходит мышечное сокращение, с помощью которого повреждение закрывается. Мышечные волокна, которые формируются вокруг раны, вначале ориентированы случайно и только постепенно приходят к ортогональному расположению. Предсуществующие мышечные волокна и продольные нейритные пучки проникают в новообразованную и служат шаблоном для дифференцировки новых миоцитов и нейронов. Развитие, регенерация и тканевой гомеостаз контролируются соматическими стволовыми клетками, известными как необласты. Необласты имеют крупное ядро и тотипотентны , то есть могут развиться в клетки любого типа. В отличие от других многоклеточных животных , у которых стволовые клетки связаны с эпидермисом, у Acoela они располагаются исключительно в паренхиме. Показано, что у Isodiametra pulchra клетки зародышевой линии и субпопуляция соматических необластов экспрессируют гомолог гена piwi , сайленсинг которого не влияет на пролиферацию клеток у зрелых особей, однако влияет на их способность к размножению и делает их потомство нежизнеспособным .
Развитие
Эмбриональное развитие Acoela начинается с дуэтного спирального дробления , которое, как считают, возникло независимо от квартетного спирального дробления, присущего всем Lophotrochozoa . Как и при квартетном спиральном дроблении, первое горизонтальное деление неравное и приводит к образованию микромеров, однако оно происходит на двуклеточной стадии, а не четырёхклеточной, поэтому микромеры образуют дуэты, а не квартеты. Первый, второй и третий дуэты микромеров дают начало всем эктодермальным структурам, а эндодермальные (паренхима) и мезодермальные (мышцы) элементы происходят от третьего дуэта макромеров. Гаструляция происходит путём нарастания микромеров поверх макромеров, и рот прорывается в месте, отличном от бластопора . Внутренние ткани появляются посредством деламинации, то есть клетки паренхимы, мышц и нервной ткани откладываются по направлению снаружи внутрь зародыша. Иногда вместо деламинации происходит миграция клеток, которые дают начало эндодерме и мезодерме. К концу гаструляции зародыш демонстрирует слоистое строение: самый верхний слой образован зачаточным эпителием, в среднем слое залегают клетки — предшественники мышц и нейронов, а самый внутренний слой сформирован клетками, которые впоследствии дадут начало пищеварительному синцитию. После гаструляции следует органогенез , при котором обособляются ресничный эпителий, субэпителиальные мышечные волокна, а на переднем конце зародыша начинает дифференцировку нервная система. Детали развития нервной системы различаются у разных семейств . Как и у остальных беспозвоночных , важную роль в нейрогенезе Acoela играют транскрипционные факторы семейства bHLH . Стадия личинки у Acoela отсутствует, то есть наблюдается прямое развитие . , описанная в 2015 году, культивировалась в лаборатории на протяжении нескольких лет. Эмбриональное развитие представителей этого вида при температуре 20 °C занимало менее двух дней .
Экология

на мадрепоровом коралле Plerogyra
Большинство бескишечных турбеллярий — это бентосные животные, обитающие на морском дне; обнаруживаются в водах всех океанов . Они могут жить под камнями, в песке или иле, а также среди водорослей. Иногда они достигают высокой плотности, например, Haplogonaria syltensis , обитающая в насыщенных водой песчаных пляжах, может достигать плотности 4000 особей на 10 см³. Черви вида Paratomella rubra могут прикрепляться к субстрату при помощи специальных прикрепительных ресничек с уплощёнными свободными концами . Зелёные черви Symsagittifera roscoffensis во время отлива держатся в верхнем слое песка, придавая ему зеленоватый оттенок. При вибрации грунта животные быстро уходят на глубину, и зеленоватый оттенок исчезает . Некоторые виды бескишечных турбеллярий пелагические , некоторые представители приспособились к жизни в пресной воде . Некоторые Acoela, например, Hofstenia miamia , обитают на погружённых в воду листьях и корнях мангров . Другие же обитают на правах комменсалов (по другому источнику, как паразиты или эндосимбионты ) в кишечниках иглокожих . Около острова Роттнест в Западнoй Австралии бескишечные из рода были обнаружены обитающими на мадрепоровых кораллах . Кроме того, представители этого рода ( Waminoa brickneri ) обитают на кораллах и в Красном море , а также вблизи Индонезии . На кораллах обитают также представители рода Convolutriloba . В образцах льда из Баренцева моря , моря Лаптевых и вод Гренландии нередко обнаруживаются ярко-красные бескишечные турбеллярии. Они населяют систему пор и щелей в кусках льда .
Большинство видов Acoela — хищники. В их рацион могут входить мелкие членистоногие , двустворчатые моллюски и различные черви , в том числе другие виды бескишечных турбеллярий. По крайней мере один вид — — практикует каннибализм . Некоторые виды потребляют одноклеточные водоросли и даже бактерии .
Систематика
Классификация
К бескишечным турбелляриям относят около 380 описанных видов (по другому источнику — около 400 ). На апрель 2018 года таксон включает 17 семейств, больше половины из которых объединены в подотряд Bursalia :
|
|
Филогенетические связи в пределах Acoela

Из-за малых размеров и небольшого количества органов, морфология которых может отличаться, классифицировать бескишечных турбеллярий, не прибегая к молекулярным методам, очень непросто. Первый вид Acoela Convoluta convoluta был описан в 1806 году и отнесён к плоским червям. Далее были описаны многие другие виды Acoela, и в 1968 году было предложено разделение этой группы на семейства в составе типа плоских червей. Это разделение производилось на основе изучения копулятивного аппарата при помощи световой микроскопии . В дальнейшем для изучения особенностей строения разных представителей Acoela применялись такие методы, как электронная микроскопия и конфокальная микроскопия в комбинации с методами и использованием меченного флуорофором фаллоидина . С помощью этих подходов удалось досконально изучить строение нервной системы, мышц стенки тела и копулятивного аппарата, а также ультраструктуру сперматозоидов .
В 2011 году группой Jondelius et al. было построено филогенетическое дерево Acoela на основании последовательностей рДНК и генов , кодирующих . Эти последовательности были известны для примерно трети видов Acoela; из-за отсутствия данных дерево не включало семейство Anthroposthiidae, а также монотипные семейства Antigonariidae, Nadinidae и Taurididae. По данным этого исследования, самым базальным семейством Acoela является Diopisthoporidae; вслед за ним идут семейство Paratomellidae и клада Prosopharyngida, состоящая из семейств Hallangidae, Hofsteniidae и Solenofilomorphidae. Базальное положение этих групп подтверждается и морфологическими данными. Эти пять семейств резко отделены от так называемых «высших» бескишечных турбеллярий, а именно, представителей клады Crucimusculata. Морфологически «высшие» Acoela характеризуются наличием вентральных поперечных мышц. Наконец, несколько семейств Acoela так и не удалось объединить в какие-либо клады .
В 2016 году с помощью метода метабаркодирования 18S рДНК было показано, что в действительности Acoela гораздо более разнообразны, чем предполагает текущая систематика. Кроме того, в пределах Acoela была выявлена клада, состоящая из обитателей бентоса, которая является сестринской по отношению ко всем остальным Acoela .
Положение Acoela в царстве животных
В 1904 году была предложена гипотеза, согласно которой животное, похожее на планулу стрекающих , дало начало предковому двусторонне-симметричному животному, схожему с современными бескишечными турбелляриями. Эта гипотеза предполагает, что примитивные бескишечные черви дали начало всем остальным двусторонне-симметричным животным. С развитием филогенетики в середине XX века появилось представление, что предок Bilateria имел целом . Acoela стали рассматриваться как вторично упрощённые плоские черви, и вместе с Nemertodermatida они были объединены в кладу Acoelomorpha в составе типа плоских червей .
Общий план строения тела и наличие статоциста сближают Acoela и Nemertodermatida, однако у последних статоцист содержит два статолита, а не один, как у Acoela. Как у Acoela, у Nemertodermatida отсутствуют органы выделения и протоки половых желёз. Однако самыми убедительными доказательствами родства этих двух групп червей является одинаковая система ресничных корешков и горизонтальная ориентация плоскости второго асимметричного деления дробления .
|
Современные представления о месте Acoela в системе Metazoa
|
Морфологический анализ, проведённый в 1986 году, и ранний кладистический анализ 1996 года указали на необходимость выделения Acoelomorpha из типа плоских червей. Изучение последовательностей рДНК показало, что Acoela находятся в самом основании клады Bilateria. Любопытно, что этот анализ разделил Acoela и Nemertodermatida. Впрочем, сравнение аминокислотных последовательностей митохондриальных белков (2004, 2010) и ( англ. Expressed sequence tag, EST ) (2009, 2011) всё-таки подтвердило близость этих двух групп и состоятельность клады Acoelomorpha. Кроме того, в дальнейшем оказалось, что гены рРНК плохо подходят для определения положения Acoela в животном царстве , так как они имеют низкий GC-состав и часто оказываются усечёнными и модифицированными. Многие исследователи также указали, что нельзя реконструировать дерево животного царства на такой небольшой выборке генов . Митохондриальные геномы тоже плохо подходят для этой цели: их последовательности неизвестны для большинства представителей Acoelomorpha, а прочитанный митохондриальный геном Symsagittifera roscoffensis оказался слишком отличающимся от митохондриальных геномов других двусторонне-симметричных животных. Для реконструкции эволюции неплохо подходят аминокислотные последовательности митохондриальных белков и EST. Анализ этих данных показал, что возможны три основных варианта филогенетического положения Acoelomorpha :
- Acoelomorpha — самая рано отделившаяся группа Bilateria;
- Acoelomorpha — сестринская группа червей Xenoturbella , и они вместе представляют собой самую рано отделившуюся группу (Xenacoelomorpha) двусторонне-симметричных животных;
- Acoelomorpha и Xenoturbella — вторичноротые животные.
В пользу последнего сценария говорит наличие специфических микроРНК у Acoelomorpha, Xenoturbella и вторичноротых, наличие белка сперматозоидов RSB66 и протеинкиназы и отсутствие перечисленных молекул у других групп животных. Впрочем, данные по микроРНК ненадёжны, потому что многие из них постоянно утрачиваются в разных группах животных, а белки RSB66 и GNE могли быть утрачены всеми первичноротыми . Однако в двух исследованиях значения бутстрэпа для группы Xenacoelomorpha оказались довольно низкими, а морфологическая схожесть этих червей может быть обусловлена одинаковым образом жизни .
Самые последние молекулярные данные указывают на то, что Xenoturbella и Acoelomorpha образуют монофилетическую группу, поэтому в настоящее время их объединяют в тип Xenacoelomorpha (к нему также относят Nemertodermatida ). Несмотря на подтверждённую монофилию Xenacoelomorpha, их точное систематическое положение — в качестве базальной группы двусторонне-симметричных животных или же базальной группы вторичноротых — не определено до сих пор. Анализ транскриптома Xenoturbella profunda показал, что Xenoturbella — сестринская группа Acoelomorpha в составе Xenacoelomorpha, а Xenacoelomorpha, в свою очередь, — сестринская группа Nephrozoa , в которую включаются первичноротые и вторичноротые животные; следовательно, по данным этого анализа Xenacoelomorpha является базальной группой двусторонне-симметричных животных .
Примечания
- ↑ , с. 439.
- ↑ Cannon J. T. , Vellutini B. C. , Smith J 3rd. , Ronquist F. , Jondelius U. , Hejnol A. (англ.) // Nature. — 2016. — 4 February ( vol. 530 , no. 7588 ). — P. 89—93 . — doi : . — .
- ↑ Отряд (англ.) в Мировом реестре морских видов ( World Register of Marine Species ). (Дата обращения: 26 мая 2018) .
- ↑ Achatz Johannes G. , Chiodin Marta , Salvenmoser Willi , Tyler Seth , Martinez Pedro. (англ.) // Organisms Diversity & Evolution. — 2012. — 29 September ( vol. 13 , no. 2 ). — P. 267—286 . — ISSN . — doi : .
- Jimenez-Guri E. , Paps J. , Garcia-Fernandez J. , Salo E. (англ.) // The International Journal Of Developmental Biology. — 2006. — Vol. 50 , no. 8 . — P. 675—679 . — doi : . — .
- Hooge M. D. , Tyler S. (англ.) // Journal of Zoological Systematics and Evolutionary Research. — 2005. — May ( vol. 43 , no. 2 ). — P. 100—113 . — ISSN . — doi : .
- , с. 412—414.
- , с. 233.
- ↑ Ya.I. Zabotin. (англ.) // Invertebrate Zoology. — 2017. — Vol. 14, no. 2 . — P. 217—225. — doi : . 21 июня 2018 года.
- , с. 232—233.
- , с. 408.
- ↑ , с. 412.
- Egger Bernhard , Steinke Dirk , Tarui Hiroshi , De Mulder Katrien , Arendt Detlev , Borgonie Gaëtan , Funayama Noriko , Gschwentner Robert , Hartenstein Volker , Hobmayer Bert , Hooge Matthew , Hrouda Martina , Ishida Sachiko , Kobayashi Chiyoko , Kuales Georg , Nishimura Osamu , Pfister Daniela , Rieger Reinhard , Salvenmoser Willi , Smith Julian , Technau Ulrich , Tyler Seth , Agata Kiyokazu , Salzburger Walter , Ladurner Peter. (англ.) // PLoS ONE. — 2009. — 11 May ( vol. 4 , no. 5 ). — P. e5502 . — ISSN . — doi : .
- Smith Julian P. S. , Tyler Seth. (англ.) // Hydrobiologia. — 1986. — January ( vol. 132 , no. 1 ). — P. 71—78 . — ISSN . — doi : .
- Gschwentner Robert , Baric Sanja , Rieger Reinhard. (англ.) // Invertebrate Biology. — 2005. — 11 May ( vol. 121 , no. 2 ). — P. 95—103 . — ISSN . — doi : .
- , с. 417—418.
- Gazizova G. R. , Zabotin Y. I. , Golubev A. I. // Morfologiia (Saint Petersburg, Russia). — 2015. — Т. 148 , № 6 . — С. 75—78 . — .
- Mieke Boone UGent, Maxime Willems, Myriam Claeys UGent, Tom Artois and Gaetan Borgonie. // Euro Evo Devo Meeting, 2nd, Abstracts. — 2008. — P. 207. 21 июня 2018 года.
- , с. 419.
- Chiodin M. , Achatz J. G. , Wanninger A. , Martinez P. (англ.) // Journal Of Experimental Zoology. Part B, Molecular And Developmental Evolution. — 2011. — 15 September ( vol. 316 , no. 6 ). — P. 427—439 . — doi : . — .
- Semmler Henrike , Bailly Xavier , Wanninger Andreas. (англ.) // Frontiers in Zoology. — 2008. — Vol. 5 , no. 1 . — P. 14 . — ISSN . — doi : .
- , с. 413—417.
- ↑ Kotikova E. A. , Raikova O. I. (англ.) // Journal of Evolutionary Biochemistry and Physiology. — 2008. — February ( vol. 44 , no. 1 ). — P. 95—108 . — ISSN . — doi : .
- , с. 415—416.
- , с. 415.
- Todt Christiane , Tyler Seth. (англ.) // Acta Zoologica. — 2006. — 15 December ( vol. 88 , no. 1 ). — P. 41—58 . — ISSN . — doi : .
- ↑ Jondelius Ulf , Wallberg Andreas , Hooge Matthew , Raikova Olga I. (англ.) // Systematic Biology. — 2011. — 9 August ( vol. 60 , no. 6 ). — P. 845—871 . — ISSN . — doi : .
- Todt Christiane. (англ.) // Journal of Morphology. — 2009. — March ( vol. 270 , no. 3 ). — P. 271—290 . — ISSN . — doi : .
- , с. 420.
- , с. 425—426.
- , с. 429—430.
- Hooge Matthew D. , Smith Julian P. S. (англ.) // Zootaxa. — 2004. — 22 February ( vol. 442 , no. 1 ). — P. 1 . — ISSN . — doi : .
- Zabotin Ya. I. , Golubev A. I. (англ.) // Biology Bulletin. — 2014. — December ( vol. 41 , no. 9 ). — P. 722—735 . — ISSN . — doi : .
- , с. 430—431.
- Petrov Anatoly , Hooge Matthew , Tyler Seth. (англ.) // Journal of Morphology. — 2006. — Vol. 267 , no. 5 . — P. 634—648 . — ISSN . — doi : .
- , с. 435.
- Petrov Anatoly , Hooge Matthew , Tyler Seth. (англ.) // Invertebrate Biology. — 2005. — 11 May ( vol. 123 , no. 3 ). — P. 183—197 . — ISSN . — doi : .
- Raikova O. I. (англ.) // Doklady Biological Sciences. — 2002. — Vol. 382 , no. 1/6 . — P. 48—50 . — ISSN . — doi : .
- ↑ Bourlat Sarah J. , Hejnol Andreas. (англ.) // Current Biology. — 2009. — April ( vol. 19 , no. 7 ). — P. R279—R280 . — ISSN . — doi : .
- Semmler H. , Bailly X. , Wanninger A. (англ.) // Frontiers In Zoology. — 2008. — 19 September ( vol. 5 ). — P. 14—14 . — doi : . — .
- ↑ , с. 430.
- Tekle Yonas I. , Raikova Olga I. , Jondelius Ulf. (англ.) // Acta Zoologica. — 2006. — 9 May ( vol. 87 , no. 2 ). — P. 121—130 . — ISSN . — doi : .
- , с. 428—429.
- Perea-Atienza E. , Sprecher S. G. , Martínez P. (англ.) // EvoDevo. — 2018. — Vol. 9 . — P. 8—8 . — doi : . — .
- Zauchner Thomas , Salvenmoser Willi , Egger Bernhard. (англ.) // Zootaxa. — 2015. — 1 April ( vol. 3941 , no. 3 ). — P. 401 . — ISSN . — doi : .
- , с. 234.
- Kolasa Jerzy , Faubel Anno. (англ.) // Bolletino di zoologia. — 1974. — January ( vol. 41 , no. 2 ). — P. 81—85 . — ISSN . — doi : .
- Cooper Crystal , Clode Peta L. , Thomson Damian P. , Stat Michael. (англ.) // Zoological Science. — 2015. — October ( vol. 32 , no. 5 ). — P. 465—473 . — ISSN . — doi : .
- Ogunlana Maxina V. , Hooge Matthew D. , Tekle Yonas I. , Benayahu Yehuda , Barneah Orit , Tyler Seth. (англ.) // Zootaxa. — 2005. — 14 June ( vol. 1008 , no. 1 ). — P. 1 . — ISSN . — doi : .
- Haapkylä Jessica , Seymour Adrian S. , Barneah Orit , Brickner Itzchak , Hennige Sebastian , Suggett David , Smith David. (англ.) // Marine Biology. — 2009. — 17 February ( vol. 156 , no. 5 ). — P. 1021—1027 . — ISSN . — doi : .
- Christine Friedrich & Jan Heldelberg. On the ecology of Acoela living in the Arctic Sea ice // Belg. J. Zool.. — 2001. — Vol. 131. — P. 213—216.
- , с. 420—425.
- Arroyo A. S. , López-Escardó D. , de Vargas C. , Ruiz-Trillo I. (англ.) // Biology Letters. — 2016. — September ( vol. 12 , no. 9 ). — doi : . — .
- ↑ Rouse G. W. , Wilson N. G. , Carvajal J. I. , Vrijenhoek R. C. // Nature . — 2016. — Vol. 530, no. 7588. — P. 94—97. — doi : . — .
- Nakano H. , Lundin K. , Bourlat S. J. , Telford M. J. , Funch P. , Nyengaard J. R. , Obst M. , Thorndyke M. C. // Nature Communications. — 2013. — Vol. 4. — P. 1537. — doi : . — .
- Hejnol A. , Obst M. , Stamatakis A. , Ott M. , Rouse G. W. , Edgecombe G. D. , Martinez P. , Baguñà J. , Bailly X. , Jondelius U. , Wiens M. , Müller W. E. , Seaver E. , Wheeler W. C. , Martindale M. Q. , Giribet G. , Dunn C. W. // Proceedings of the Royal Society. Ser. B. Biological Sciences. — 2009. — Vol. 276, no. 1677. — P. 4261—4270. — doi : . — .
Литература
- Рупперт Э. Э., Фокс Р. С., Барнс Р. Д. Протисты и низшие многоклеточные // Зоология беспозвоночных: функциональные и эволюционные аспекты. — М. : Издательский центр «Академия», 2008. — Т. 1. — 496 с.
- Зоология беспозвоночных : в 2 т. / под ред. В. Вестхайде и Р. Ригера . — М. : Товарищество научных изданий КМК, 2008. — Т. 1: от простейших до моллюсков и артропод. — 512, [8] с. — ISBN 978-5-87317-491-1 .
|
|
|
|---|---|
| Таксономия |
|
| В библиографических каталогах |
|
- 2020-04-22
- 1

