Слепни
- 1 year ago
- 0
- 0
Микроспори́дии ( лат. Microsporidia ) — клада родственных грибам простейших , все представители которой являются облигатными внутриклеточными паразитами эукариотических организмов. Описано около 1300 видов в 160 родах, что является малой частью реального разнообразия данной группы, так как огромное количество потенциальных хозяев не было исследовано на предмет заражения микроспоридиями . Данные патогены широко распространены среди животных практически всех систематических групп, от простейших до высших позвоночных , включая человека. Наиболее многочисленны и разнообразны микроспоридии ракообразных и насекомых .
Микроспоридии обладают рядом уникальных признаков, которые демонстрирует крайнюю степень специализированности данного таксона к внутриклеточному паразитизму. Споры данных патогенов содержат уникальный комплекс органелл, известный только для микроспоридий — аппарат экструзии , предназначенный для заражения клетки хозяина путём прокола её оболочки и вбрасывания зародыша непосредственно в цитоплазму . Такой способ переноса зародыша из споры в клетку животного-хозяина не известен для других простейших . Геном микроспоридий является наименьшим среди эукариот и в нём практически отсутствуют интроны , а митоз носит форму закрытого внутриядерного плевромитоза. Их рибосомы схожи с прокариотическими и имеют коэффициент седиментации 70S. Клетка микроспоридий лишена кинетосом и их производных ( жгутиков и центриолей ), лизосом , гранул запасных питательных веществ . Долгое время считалось первичным отсутствие у микроспоридий митохондрий , но в недавнем времени в их клетках были обнаружены крохотные двумембранные органеллы — митосомы и было доказано их митохондриальное происхождение .
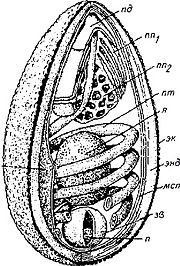
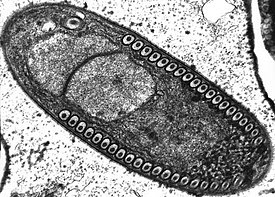
Оболочка споры у подавляющего большинства видов микроспоридий состоит из трёх слоёв: гликопротеиновой экзоспоры, хитиновой эндоспоры и цитоплазматической мембраны. Экзоспора может быть многослойной и формировать придатки различной формы .
Аппарат экструзии включает в себя заднюю вакуоль , якорный диск , поляропласт и полярную трубку . Задняя вакуоль представлена одной или несколькими камерами, расположенными терминально. Иногда внутри вакуоли бывает расположена постеросома — рудимент аппарата Гольджи . Во время экструзии споры данная органелла резко увеличивается в размере, выталкивая зародыш в полярную трубку >.
Поляропласт чаще всего представляет собой упаковку плотно уложенных мембран, но также может включать различные везикулы и трубчатые структуры. Считается, что поляропласт задействован в создании высокого внутриспорового давления, необходимого для выворачивания полярной трубки, а также является «поставщиком мембран» для самой трубки и проходящей по ней спороплазмы .
Полярная трубка представлена удлинённой двумембранной структурой с умеренно электроноплотным содержимым, отходящей от расположенного на переднем конце споры якорного диска и уложенной спирально. Длина, степень развития и форма полярных трубок у микроспоридий широко варьируют. Обычно, число витков спирали колеблется от 6 до 12, но у некоторых видов достигает 36. Представители семейства Metchnikovellidae — паразиты других одноклеточных организмов (тоже паразитических) грегарин имеют крайне редуцированный аппарат экструзии, с полярной трубкой по размерам не превышающей длину споры. По особенностям строения выделяют несколько типов полярных трубок: изофилярная — обладает одинаковым размером на всём протяжении; анизофилярная — диаметр первых витков значительно превышает диаметр последующих; гетерофилярная — различные витки имеют различный диаметр. После выброса полярная трубка становится полой, и через неё происходит внедрение спороплазмы в клетку хозяина .
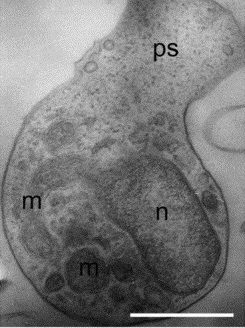
Спороплазма (зародыш) представляет собой лежащее в споре одиночное ядро или диплокарион, окружённое тонким слоем цитоплазмы с рибосомами. После экструзии споры зародыш проходит через полярную трубку и оказывается в цитоплазме хозяина. При этом в заражённой клетке спороплазма оказывается окружённой собственной цитоплазматической мембраной, происхождение которой до сих пор доподлинно не известно .
Пролиферативная стадия — меронт — представлена интенсивно растущей и размножающейся клеткой, одноядерной или многоядерной, в зависимости от стадии . Цитоплазма меронтов содержит минимальный набор органелл: аппарат Гольджи трубчатого типа, строение которого уникально для микроспоридий , рибосомы, гладкий и шероховатый эндоплазматический ретикулум . Кроме того, в цитоплазме меронтов можно найти различные трубчатые структуры и везикулы .
При переходе к первой стадии спорогонии — споронту — клетка микроспоридии приобретает поверх цитоплазматической мембраны дополнительную оболочку, первое время заметную только при электронном микроскопировании. На этом этапе продолжается деление ядер, в результате чего образуются многоядерные спорогональные плазмодии , дающие начало споробластам .
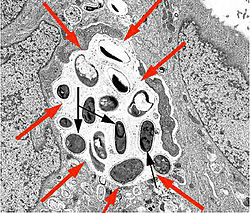
У многих видов микроспоридий снаружи от оболочки споронта закладывается ещё один покровный слой, образующий оболочку спорофорного пузырька (по старой терминологии панспоробласта ). Строение спорофорных пузырьков и развитие полости между оболочками широко варьирует у разных видов микроспоридий .
Споробласт — переходная стадия между спорогональным плазмодием и спорой. Характеризуется интенсивным развитием оболочек и закладкой всех будущих органелл споры .
Геном микроспоридий, как и у большинства других эукариот, состоит из набора линейных хромосом , но является, однако, крайне редуцированным и необычным. Средний размер геномов микроспоридий чрезвычайно мал, у Encephalitozoon intestinalis он составляет всего 2,3 миллиона пар нуклеотидов , что даже меньше, чем у некоторых бактерий . Набор кодируемых белков также невелик, в геноме найдено около 2000 белоккодирующих последовательностей. Геном микроспоридий сильно компактизирован, и в нём практически отсутствуют интроны , при этом гены , гомологичные генам других организмов, у микроспоридий часто бывают значительно короче .
В процессе адаптации к внутриклеточному паразитизму микроспоридии утратили большинство метаболических путей, таких как окислительное фосфорилирование или цикл Кребса . На стадии внутриклеточного развития паразит вообще не использует собственную систему энергетического метаболизма , полностью полагаясь на транспорт АТФ из цитоплазмы клетки хозяина за счёт уникальных АТФ-переносчиков . Данные переносчики пластидно-бактериального типа были приобретены микроспоридиями за счёт горизонтального переноса генов от бактерий и являются уникальными для данного таксона. Таким образом, микроспоридии дальше всех других эукариот продвинулись на пути адаптации к внутриклеточному паразитизму, так как кроме них пока не было найдено паразитов, способных напрямую транспортировать АТФ из хозяйской клетки .
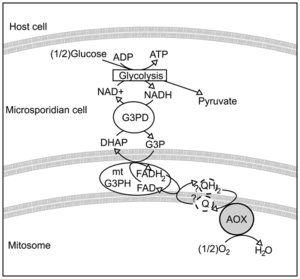
На стадии споры микроспоридии поддерживают свою жизнедеятельность за счёт гликолиза . При этом, долгое время было неясно, каким образом происходит реокисление образующихся в ходе гликолиза восстановительных эквивалентов, таких как NADH . На сегодняшний день предлагается следующая схема данного процесса, включающая и альтернативную оксидазу . В цитоплазме цитоплазматическая форма глицерол-3-фосфатдегидрогеназы окисляет NADH до NAD + , восстанавливая при этом дигидроксиацетонфосфат до глицерол-3-фосфата. Последний поступает в межмембранное пространство митосомы , где вновь окисляется до дигидроксиацетонфосфата за счёт FAD -зависимой митосомальной формы глицерол-3-фосфатдегидрогеназы. Восстановленная форма FADH2, в свою очередь, передаёт электроны на пул убихинонов , расположенный во внутренней мембране митосомы. Конечным этапом является реокисление убихинона ферментом альтернативная оксидаза с использованием кислорода в качестве конечного акцептора электронов и восстановлением последнего до воды . В геномах представителей группы Terresporidia (включает в себя большинство микроспоридий, паразитирующих в наземных животных) гена альтернативной оксидазы найдено не было. Таким образом, схема энергетического обмена у этой группы до сих пор остаётся загадкой .

Инвазионной стадией у микроспоридий является спора. Она содержит зародыш (спороплазму) и сложно организованный аппарат экструзии, обеспечивающий выброс зародыша из споры и внедрение его в клетку хозяина. Для многоклеточных животных заражение обычно происходит при попадании спор паразитов в пищеварительный тракт хозяина. В отличие от других внутриклеточных паразитов, в подавляющем большинстве случаев развитие микроспоридий происходит в прямом контакте с цитоплазмой клетки хозяина, без образования паразитофорной вакуоли . После внедрения происходит рост и дифференциация зародыша, после чего клетка паразита превращается в меронт, пролиферативную стадию, которая размножается бинарным или множественным делением по типу . Затем микроспоридии приступают к спорогонии. Ядра споронтов претерпевают от одного до нескольких делений, в результате чего образуются спорогональные плазмодии, дающие начало споробластам, в которых происходит формирование спор. Обычно на этом этапе происходит разрушение заражённой клетки хозяина, а вышедшие таким образом споры заражают другие клетки того же организма или выводятся наружу для заражения других особей.
Микроспоридии демонстрируют очень высокую степень разнообразия жизненных циклов . Описанный выше моноксенный (включающий только одного хозяина) цикл с образованием спор одного типа характерен для почти 80 % описанных видов . Однако и в таком цикле особенности спорогонии, строение ядерного аппарата, количество и типы делений на всех стадиях широко варьируют. В остальных случаях цикл может проходить более чем в одном хозяине, включать несколько спорогоний с образованием нескольких типов спор, различающихся морфологически и функционально. Для нескольких видов на различных стадиях развития описан мейоз и половой процесс, проходящий по типу гаметоцитогамии . Примером наиболее сложных поликсенных жизненных циклов, включающих половой процесс и морфо-функциональную дифференциацию спор, являются жизненные циклы микроспоридий родов Amblyospora и Parathelonahia .
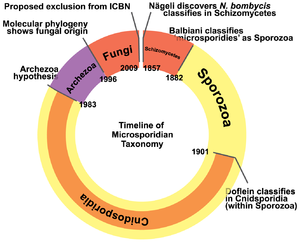
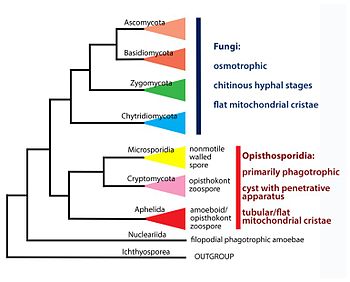
С момента первого описания в 1857 году положение таксона в системе живой природы постоянно менялось. Первоначально микроспоридии были отнесены к группе Schizomycetes , куда входили различные виды дрожжей и бактерий. Затем, в начале XX века таксон был вместе с другими внутриклеточными паразитами включен в состав группы Sporozoa в подгруппу , включающую также миксоспоридий , актиноспоридий и хеликоспоридий. Во второй половине XX века было накоплено достаточно данных, доказывающих искусственность группы Sporozoa , и микроспоридии были включены в группировку , объединяющую одноклеточных организмов без митохондрий. Предполагалось, что представители данной группы являются наиболее древними из современных эукариот и отделились от их общего предка ещё до приобретения митохондрий. Несмотря на то, что сейчас можно считать доказанными ошибочность гипотезы Archezoa и вторичность потери микроспоридиями митохондрий, часть молекулярных исследований, выполненных в конце XX века, поддерживали данную гипотезу . Однако, после увеличения числа исследуемых генов, все больше молекулярно- филогенетических исследований демонстрировали родство микроспоридий с грибами. При этом в разных работах положение микроспоридий относительно грибов оказывается различным. По результатам некоторых исследований микроспоридии входят в состав грибов , однако последние работы обычно выявляют микроспоридий, как сестринскую грибам группу . На сегодняшний день принято формальное включение микроспоридий в таксон Fungi , однако такая точка зрения подвергается критике со стороны многих специалистов , и вполне вероятно, что систематическое положение микроспоридий ещё не раз кардинально изменится.
Классификация микроспоридий подвергается непрерывным перестройкам на основе использования новых таксономических признаков, полученных при изучении тонкого строения, жизненных циклов , цитохимических и молекулярно-биологических свойств как новых, так и известных форм. В классической системе, построенной на морфологических признаках, общепризнано разделение типа на два класса, один из которых включает формы микроспоридий, образующие в конце коротких жизненных циклов споры с примитивным аппаратом экструзии, а другой — более сложно организованные формы с моно- или диксенными жизненными циклами, завершающимися образованием одного-двух-трёх типов спор с хорошо развитым аппаратом экструзии . При подразделении на отряды и семейства основное внимание уделено строению, в основном, стадий спорогонии, типу ядерного аппарата, хромосомным и жизненным циклам. Однако анализ нуклеотидных последовательностей рибосомальной РНК показывает отсутствие связи между классическими признаками морфологии и филогенетическим родством между видами микроспоридий, что требует пересмотра значения морфологических критериев, используемых в систематике. Использование подходов молекулярной филогении позволяет разделить проанализированные формы микроспоридий на пять филогенетических ветвей, объединяемых в три класса, исходя из первичной приуроченности паразитов к месту обитания хозяев .

Микроспоридии вызывают хозяйственные потери на шелководнях и пасеках, будучи возбудителями эпизоотий нозематозов у медоносных пчёл ( Apis mellifera ), тутового шелкопряда ( Bombyx mori ). Описаны случаи массовой гибели других полезных беспозвоночных животных, например, мидий Mytilus edulus , а также различных промысловых видов рыб .
Особую проблему представляют микроспоридиозы человека. Обычно микроспоридии вызывают тяжелые и быстро развивающиеся оппортунистические инфекции , возникающие на фоне иммунодефицита . Однако данные патогены могут быть опасны и для иммунокомпетентных людей; было показано, что микроспоридия Encephalitozoon cuniculi , паразитирующая у кроликов, способна заразить человека с нормальной иммунной системой . Клинические проявления микроспоридиоза очень разнообразны и варьируют в зависимости от вида микроспоридии-возбудителя инфекции . Наиболее часто встречаемым симптомом является диарея .
Микроспоридии, паразитирующие у насекомых -вредителей сельскохозяйственного, медицинского и ветеринарного значения, являются перспективными агентами биологической борьбы с этими видами. Некоторые виды микроспоридий комаров вызывают высокую смертность личинок, снижают плодовитость самок, а также предотвращают развитие нескольких видов малярийных плазмодиев . В 1978 году в США для борьбы с прямокрылыми вредителями был создан первый промышленный биопрепарат на основе микроспоридий — «Нолок», который получил высокую экономическую оценку благодаря низкой стоимости, высокой эффективности и отсутствию негативного влияния на окружающую среду .