Interested Article - Фибрилларин

- 2020-06-20
- 1
Локус
:
![]()
Фибриллари́н ( англ. Fibrillarin, Fbl , другие названия ) — ядрышковый белок , у человека кодируется геном FBL , локализованным на 19-й хромосоме . Основной функцией фибрилларина является участие в транскрипции и процессинге рРНК , поэтому он часто используется как маркер активных ядрышек. Фибрилларин является , то есть переносит метильную группу с S-аденозилметионина на 2'- гидроксильную группу рибозы в молекуле РНК -мишени. Кроме того, фибрилларин опосредует метилирование остатка глутамина Gln105 в . Белки, по структуре и функциям соответствующие фибрилларину, были найдены и у архей , которые являются прокариотами и не имеют ядрышек. Повышенный уровень фибрилларина обнаруживается в случае многих раковых заболеваний. Кроме того, фибрилларин может взаимодействовать со многими вирусными белками .
Названия
Название «фибрилларин» используется для обозначения нескольких белков у разных организмов , в том числе архей, хотя их фибрилларин сильно отличается от фибрилларина эукариот . В литературе фигурирует множество синонимов названия «фибрилларин», в том числе ядрышковый антиген склеродермы массой 34 кДа ( англ. 34 kDa nucleolar scleroderma antigen ), Dmel_CG9888, CG9888, Dmel\CG9888ri, GCR-6, GCR6, Pen59C5, fib, pen59C5, Fib, FIB, FBL, Fbl, FIB1, FLRN, RNU3IP1, фибрилларин рРНК 2'-O-метилтрансфераза ( англ. rRNA 2'-O-methyltransferase fibrillarin ), NOP1, nop1, fibM, afib. Для обозначения фибрилларина дрожжей часто используется название NOP1, а для фибрилларинов архей — aFIBs .
Распространение

Впервые фибрилларин был описан в ядрышке слизевика Physarum polycephalum в 1977 году. Фибрилларин обнаружен не только у эукариот, но и у архей, и, поскольку археи не имеют ядрышка, предполагается, что функции фибрилларина архей несколько отличны от функций фибрилларина эукариот. В клетках дрожжей Saccharomyces cerevisiae содержащийся там фибрилларин NOP1 можно без ущерба для этих клеток заменить на фибрилларин растения Arabidopsis thaliana , лягушки Xenopus laevis или человека, но не на фибрилларин инфузории Tetrahymena thermophila (возможно, это обусловлено отличающейся структурой N-концевого домена последнего). Приведённые данные свидетельствуют в пользу высокой консервативности структуры и функций фибрилларина. Однако домен GAR, обогащённый аргинином и глицином , имеет низкую консервативность аминокислотного состава, а при замене NOP1 фибрилларином человека, X. laevis или A. thaliana всё-таки изменяются параметры ядрышка и рост дрожжевых клеток , поэтому некоторые функции фибрилларина у разных организмов, по-видимому, несколько различаются .
Консервативность аминокислотных последовательностей фибрилларина различна в разных группах организмов. Наибольшую консервативность демонстрируют фибрилларины растений (63 % сходства между наиболее удалёнными представителями группы) и позвоночных (61 %), в то время как наименее консервативны фибрилларины беспозвоночных , грибов и архей (33, 27 и 20 % соответственно) .
На основании филогенетических данных можно выделить несколько семейств фибрилларинов, однако пока для такого разделения недостаточно биохимических и генетических данных .
У человека ген FBL , кодирующий фибрилларин, находится на 19-й хромосоме в локусе 19q13.1 и содержит 9 экзонов . Ген располагается на минус-цепи и имеет длину 11962 пар оснований .
Структура
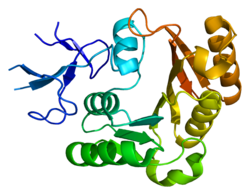
Фибрилларин принадлежит к семейству SAM-метилтрансфераз, содержащих укладку Россмана . В зависимости от организма молекулярная масса фибрилларина может составлять от 34 до 38 кДа . У человека фибрилларин состоит из 321 аминокислотного остатка и имеет массу 33784 Да . Фибрилларин можно подразделить на N-концевой домен и домен, обладающий метилтрансферазной активностью. У Arabidopsis thaliana N-концевой домен подразделяется на два участка: домен, обогащённый глицином и аргинином (GAR) и состоящий из 77 аминокислотных остатков, и спейсерный участок, включающий 61 аминокислотный остаток. Домен GAR участвует во взаимодействии с различными клеточными и вирусными белками, а также содержит сигнал, удерживающий белок в ядрышке совместно с РНК-связывающим мотивом . GAR не является необходимым доменом для локализации фибрилларина в тельцах Кахаля , однако у человека и Arabidopsis thaliana он совершенно необходим для локализации белка в ядре. Домен GAR может метилироваться по нескольким остаткам аргинина. Фибрилларин может метилироваться по остаткам аргинина белком , причём метилированные остатки составляют 45 % всех остатков аргинина фибрилларина. Метилирование может способствовать специфичному связыванию фибрилларина с некоторыми белками, например, . У человека домен GAR и спейсерный участок взаимодействуют с белком SF2A-p32, а метилтрансферазный домен взаимодействует с .
Метилтрансферазный домен подразделяется на два участка: R-, или центральный участок, включающий 87 аминокислотных остатков, и участок длиной 95 аминокислотных остатков, обогащённый α-спиралями . Для связывания фибрилларина с РНК необходимо несколько участков. Во-первых, это мотив GCVYAVCF, расположенный внутри R-участка, во-вторых, последовательность в R-участке между аминокислотными остатками 138 и 179 и, в-третьих, последовательность между аминокислотными остатками 225 и 281, локализованная в обогащённом α-спиралями участке. Два последних сайта связывания РНК взаимодействуют с различными РНК независимо друг от друга и делеция одного из них не оказывает негативного влияния на способность фибрилларина связывать РНК, однако при совместной работе этих двух сайтов проявляется синергический эффект. В С-концевом участке белка имеется консервативная структура, состоящая из 7 α-спиралей и 7 . Кроме того, в С-концевом участке находится сайт связывания S-аденозилметионина, окружённый тремя консервативными аминокислотными остатками. Этот же регион взаимодействует с белком Nop56 .
Удивительно, но структура фибрилларина высококонсервативна в ряду организмов от архей до человека. Однако фибрилларины архей лишены домена GAR, а потому короче фибрилларинов эукариот .
Как отмечалось выше, из посттрансляционных модификаций фибрилларин может подвергаться метилироваться по остаткам аргинина. Кроме того, он может фосфорилироваться по остатку серина Ser124 .
Внутриклеточная локализация
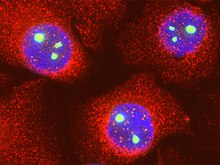
Как и многие другие ядерные белки, фибрилларин — очень динамичный белок, вероятно, из-за того, что процесс образования рибосомных частиц, в котором он принимает участие, требует высокой подвижности. Фибрилларин может быстро перемещаться между ядрышками и нуклеоплазмой , а также появляться в тельцах Кахаля. Возможно, фибрилларин активно перемещается по ядру в поисках субстрата для метилирования .
В интерфазе клеточного цикла фибрилларин локализован в плотном фибриллярном компоненте ядрышка, и при переходе от G1 - к G2-фазе его концентрация может удваиваться. В начале профазы ядрышки начинают разбираться, и фибрилларин совместно с другими элементами процессинга рРНК (пре-рРНК, нуклеолин , малые ядрышковые РНК и ) перемещается на периферию хромосом, где составляет часть перихромосомной оболочки или перихромосомного компартмента. Кроме того, фибрилларин был отмечен в цитоплазме клеток, проходящих митоз , поэтому, возможно, во время митоза комплексы процессинга рРНК разбираются и разрушаются в цитоплазме. В телофазе фибрилларин уже собирается в комплексы процессинга рРНК в пренуклеолярных тельцах (PNB). После этого PNB связываются с ядрышковыми организаторами (NORs), причём фибрилларин является одним из первых факторов процессинга рРНК, которые переходят из PNBs в NORs. Было высказано предположение, что киназы и фосфатазы , регулирующие переход из митоза в интерфазу , могут также регулировать переход фибрилларина в NORs до начала транскрипции генов рРНК (р ДНК ) .
Функции
Фибрилларин непосредственно участвует в пост-транскрипционном процессинге рРНК — в частности, разрезании пре-рРНК, метилировании рРНК и сборке рибосом. Метилирование рРНК происходит более чем в 100 сайтах с небольшими отклонениями в зависимости от организма. У архей метилирующий комплекс состоит из белков aFIB, Nop5 и L7Ae. У эукариот белки и заменяют Nop5, а белок 15.5K замещает L7Ae. N-концевой домен Nop5 взаимодействует с aFIB, а C-концевой домен Nop5 связывается с L7Ae, и при связывании с направляющей РНК комплекс становится активным. У эукариот в роли направляющих РНК выступают малые ядрышковые РНК. В метилировании рРНК участвуют малые ядрышковые РНК, содержащие C/D-бокс. С-бокс, представляющий собой последовательность RUGAUGA (R — пурин ), находится рядом с 5'-концом РНК, а D-бокс (CUGA) находится вблизи 3'-конца. Направляющая малая ядрышковая РНК содержит последовательность из 10—21 нуклеотидов , комплементарных РНК-мишени, и метилирование происходит на 5 нуклеотидов выше D-бокса. С фибрилларином взаимодействуют следующие малые РНК: U3, U8, U13, U14, U60, x, y, snR3, snR4, snR8, snR9, snR10, snR11, snR30, snR189 и snR190. Из всех многочисленных метилирований, происходящих при процессинге рРНК, по-видимому, ни одно не отвечает за какую-либо специализированную функцию. Комплекс эукариотического фибрилларина с Nop56, Nop58, 15.5K и малыми ядрышковыми РНК имеет массу около 400 кДа. Фибрилларин может входить в его состав в виде тетрамера , который меняет свою конформацию для того, чтобы метилировать различные участки рРНК. Nop56/58 играет важную роль в расположении каталитической субъединицы на РНК-мишени, одновременно взаимодействуя и с фибрилларином. Малые ядрышковые РНК выступают в роли направляющих и определяют непосредственный сайт редактирования .
Было показано, что фибрилларин может участвовать в метилировании гистона H2A по остатку глутамина Gln105 у дрожжей и Gln104 у человека. Такая модификация происходит только в ядрышке, где наблюдается высокая концентрация фибрилларина. Это — первая эпигенетическая модификация гистонов, характерная только для ядрышка. Данная модификация препятствует связыванию комплекса ремоделирования хроматина и характерна для локуса 35S рДНК, а потому оказывает эпигенетический эффект на промоторы .
Роль метилирования, опосредуемого фибрилларином, исследовалась на многоклеточных организмах (на мышах ) при помощи нокдауна соответствующего гена. Фибрилларин дикого типа заменялся формой, лишённой метилтрансферазного и N-концевого доменов и, таким образом, состоящей из одного только домена GAR. Гомозиготные по этой форме эмбрионы не развивались из-за многочисленных апоптозов , однако гетерозиготы не имели каких-либо отклонений от нормы. Кроме того, было показано, что фибрилларин необходим для нормального развития данио-рерио , а у растений сокращение экспрессии фибрилларина при помощи РНК-интерференции приводило к развитию карликового фенотипа .
Взаимодействия с другими белками
Взаимодействия с клеточными белками
Помимо вышеупомянутых белков Nop56, Nop58 и 15.5K, которые совместно с фибрилларином принимают участие в процессинге рРНК, фибрилларин взаимодействует со многими другими клеточными белками. Так, белки p32 и Nop52 взаимодействуют с фибрилларином не одновременно, но, предположительно, в одних и тех же сайтах. p32 участвует в регуляции сплайсинга , а Nop52 задействован в поздних стадиях образования рибосомных субъединиц 60S. Поскольку оба белка не могут взаимодействовать с фибрилларином одновременно, вероятно, взаимодействие каждого из них с фибрилларином необходимо на определённом этапе биогенеза рибосом. Фибрилларин может взаимодействовать с DDX5 . В тельцах Кахаля фибрилларин может взаимодействовать с белком выживания мотонейронов ( англ. Survival of Motor Neuron protein (SMN) ). У растений фибрилларин 2 является частью медиаторного комплекса транскрипции, опосредованной РНК-полимеразой II , и может взаимодействовать с основными транскрипционными факторами РНК-полимеразы II, такими как .
Взаимодействия с вирусными белками
Некоторые вирусы , в жизненных циклах которых имеется ядерная фаза ( , , ВИЧ и другие) взаимодействуют с белками, локализованными в тельцах Кахаля и ядрышках, при реализации своих репликации и транскрипции. К числу таких белков относится и фибрилларин, перемещающийся между ядрышками и тельцами Кахаля. Например, фибрилларин взаимодействует с белком ORF3, кодируемым вирусом из группы умбравирусов, поражающих растения. Фибрилларин непосредственно взаимодействует своим обогащённым аргинином доменом с обогащённым лизином доменом ORF3, благодаря чему вирусный белок может перемещаться между ядрышком и тельцем Кахаля. Кроме того, при вирусной инфекции фибрилларин переносится в цитоплазму, где участвует в образовании вирусных частиц .
С фибрилларином взаимодействуют и некоторые вирусы животных . Например, многофункциональный белок NS1 подавляет процессинг мРНК клетки-хозяина и препятствует развитию антивирусного ответа. NS1 связывается с фибрилларином и нуклеолином через свой С-концевой сигнал ядерной/ядрышковой локализации . Предполагается, что белок Tat ВИЧ взаимодействует с фибрилларином в комплексе с малой ядрышковой РНК U3 и тем самым нарушает процесс созревания пре-рРНК в ядрышках. Это, в свою очередь, нарушает жизнедеятельность поражённых клеток и может приводить к апоптозу. У других вирусов, например, у вызывающего свинку , в ходе жизненного цикла с фибрилларином взаимодействуют белки нуклео капсида . По-видимому, фибрилларин играет решающую роль в жизненном цикле генипавируса — высокопатогенного вируса, переносимого летучими мышами . В клетках, лишённых фибрилларина, наблюдались существенные нарушения синтеза РНК и белков этого вируса .
Клиническое значение
Как и другие ядерные белки, фибрилларин принимает участие в регуляции процессов деления и роста клеток, поэтому нередко нарушения его экспрессии приводят к развитию раковых заболеваний. Фибрилларин является онкогеном . Сверхэкспрессия фибрилларина наблюдается в случае простаты у мышей у человека, которая может развиться в рак простаты . В случае человеческой аденокарциномы количество фибрилларина связано с количеством белка Myc — хорошо изученного онкогена, который может взаимодействовать с фибрилларином. Превышение экспрессии фибрилларина в два раза было выявлено в клетках лейкемии и лимфомы различных типов. Показано, что p53 снижает экспрессию фибрилларина, взаимодействуя с последовательностью интрона 1 в гене фибрилларина. В клетках рака молочной железы снижение содержания p53 приводит к повышенному уровню экспрессии фибрилларина, а это, в свою очередь, увеличивает количество неправильно метилированных рРНК. Это нарушает работу рибосомы и увеличивает экспрессию онкогенов через IRES .
Аутоантитела к фибрилларину обнаруживаются примерно у 8 % пациентов, страдающих от системной склеродермии .
Примечания
- ↑ . Дата обращения: 3 октября 2017. 5 мая 2016 года.
- ↑ Rodriguez-Corona U. , Sobol M. , Rodriguez-Zapata L. C. , Hozak P. , Castano E. (англ.) // Biology of the cell / under the auspices of the European Cell Biology Organization. — 2015. — Vol. 107, no. 6 . — P. 159—174. — doi : . — .
- ↑ . Дата обращения: 21 марта 2016. 15 марта 2016 года.
- . Дата обращения: 21 марта 2016. 17 августа 2017 года.
- Nicol S. M. , Causevic M. , Prescott A. R. , Fuller-Pace F. V. (англ.) // Experimental cell research. — 2000. — Vol. 257, no. 2 . — P. 272—280. — doi : . — .
- Deffrasnes C. , Marsh G. A. , Foo C. H. , Rootes C. L. , Gould C. M. , Grusovin J. , Monaghan P. , Lo M. K. , Tompkins S. M. , Adams T. E. , Lowenthal J. W. , Simpson K. J. , Stewart C. R. , Bean A. G. , Wang L. F. (англ.) // PLoS pathogens. — 2016. — Vol. 12, no. 3 . — P. e1005478. — doi : . — .
- Peterson L. K. , Jaskowski T. D. , Mayes M. D. , Tebo A. E. (англ.) // Immunologic research. — 2016. — Vol. 64, no. 2 . — P. 483—488. — doi : . — .
- Wielosz E. , Dryglewska M. , Majdan M. (англ.) // Postepy higieny i medycyny doswiadczalnej (Online). — 2014. — Vol. 68. — P. 987—991. — doi : . — .
Литература
- / O'Day, Danton H, Catalano, Andrew. — Netherlands: Springer, 2013. — ISBN 978-94-007-5818-6 . — doi : .
- / Mark O. J. Olson. — New York: Springer, 2011. — ISBN 978-1-4614-0514-6 . — doi : .
- 2020-06-20
- 1