Interested Article - Эмбриональная диапауза

- 2020-12-31
- 1
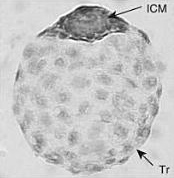
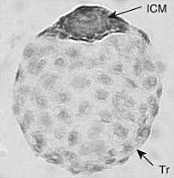
Эмбриональная диапауза ( др.-греч. ἔμβρυον — зародыш и διάπαυσις — перерыв) — феномен глубокой задержки развития эмбриона , происходящей при нормальном ходе его развития . От вида к виду стадия онтогенеза , на которой происходит задержка, сильно различается. Внешние условия влияют на эмбриональную диапаузу опосредованно; она не вызывается непосредственно неблагоприятными внешними воздействиями. . У плацентарных этот процесс чаще называют задержанной имплантацией .
Обнаружена у видов, принадлежащих к несвязанным таксономическим группам — от насекомых и до млекопитающих : у коловраток , низших ракообразных , саранчовых , тутового шелкопряда , у птиц и рыб , у около сотни видов млекопитающих , относящихся к разным отрядам , например, у грызунов и хищных . У растений есть схожий механизм, который называется .
Может происходить на разных стадиях развития эмбриона. Так, для насекомых различают ранне-, средне- и позднеэмбриональную диапаузу. У подавляющего большинства млекопитающих диапауза происходит только на одном этапе развития — на стадии бластоцисты перед имплантацией в стенку матки .
Определяющим свойством эмбриональной диапаузы у растений и животных является кардинальное уменьшение числа или прекращение митозов (разновидности клеточного деления) в эмбрионе, которое может происходить на фазе G0/G1 или G2 клеточного цикла . Возобновление деления означает прекращение диапаузы.
Эволюционное значение
Особенности протекания эмбриональной диапаузы существенно различаются от вида к виду, что даёт основания утверждать, что её распространённость вызвана не только филогенетическими связями, но и независимым возникновением у разных видов в ходе эволюции, по причине того, что в некоторых экологических ситуациях она даёт серьёзные селективное преимущество . Природа этого преимущества для большинства таксонов не выявлена, и ещё меньше известно об эволюционных связях механизма диапаузы среди современных видов.
Как эволюционный механизм эмбриональная диапауза позволяет разнести по времени процессы оплодотворения и активного развития эмбриона. Это даёт возможность производить оба процесса в наиболее благоприятные для данного вида моменты, что повышает выживаемость потомства , вплоть до того, что у некоторых видов зародыши в состоянии эмбриональной диапаузы выживают в условиях, в которых взрослые особи существовать не способны — например, у рыб и рачков в полностью высыхающих водоёмах или пройдя через желудочно-кишечный тракт хищника :270 .
История изучения
По всей видимости, первое упоминание задержанной имплантации (эмбриональной диапаузы у млекопитающих) сделано в середине первой декады 17-го века в полевых заметках Уильяма Гарвея , написанных при сопровождении короля Англии Карла I в его поездке, связанной с охотой на косуль . Считается, что первое научное описание задержанной имплантации дал Бишофф ( Bischoff ) в 1854 году , изучая европейских косуль . Иногда приоритет признаётся за Циглером ( Ziegler ), 1843 год .
Эмбриональная диапауза у насекомых изучалась вместе с другими видами диапаузы (личиночной и имажинальной).
Термин « диапауза » был введён Уилером в 1893 году для описания некоторых состояний при развитии эмбриона длинноусых прямокрылых ( Xiphidium ensiferum ). В его работе под диапаузой понималась стадия покоя между разнонаправленными движениями эмбриона в желтке. Новое значение термину дал Геннеги ( Henneguy ) в статье 1904 года; в ней под диапаузой понималось состояние остановки развития как эмбриона, так и взрослого насекомого. Выделение двух типов диапаузы — факультативной и облигатной — было предложено Стейнбергом ( Steinberg ) и Каменски ( Kamensky ) в 1936 году.
Для эмбриональной паузы у членистоногих часто используется термин «яйцевая диапауза» ( англ. egg diapause , ovarian diapause ). Предположения о том, что эмбриональная диапауза может быть связана с гормонами, встречаются в работах Копеца ( Kopec ) 1917-го и 1922-го годов. В 1924 японский исследователь Ватанабе ( Watanabe ) пришёл к выводу, что диапауза у яиц шелкопряда связана с получением «ингибирующего» вещества от материнского мотылька. Изучение связанных с эмбриональной диапаузой гормональных процессов тутового шелкопряда ( Bombyx mori ) связано с именами следующих исследователей: Фукуда ( Fukuda ), Кинсаку Хасэгава ( Kinsaku Hasegawa ), Лис ( Lees ) и де Вилд ( de Wilde ). Фукуда и Хасэгава независимо друг от друга 1951 году сообщили выводы о нейрогормональном механизме управления диапаузой. В начале 1990-х структура «гормона диапаузы» была выявлена . Однако, по всей видимости, этот механизм диапаузы не является универсальным для насекомых .
Термин «эмбриональная диапауза» ( англ. embryonic diapause ) был предложен австралийским зоологом Сесилем Хью Тиндэйл-Биско ( Cecil Hugh Tyndale-Biscoe ) в 1963 году в работе «Роль жёлтого тела в задержанной имплантации у сумчатых » . Его предложение основывалось на том, что у сумчатых возобновившая развитие бластоциста , в отличие от плацентарных млекопитающих, имплантируется в стенку матки не сразу, в связи с чем ранее использованный термин «задержанная имплантация» ( англ. delayed implantation ) неточно описывал процесс. . Позже термин был распространён на всех млекопитающих и другие организмы.
Дискуссия о разнообразии проявлений задержанной имплантации у близкородственных видов в 1980-х связывается с именами Микаэля Санделла ( Mikael Sandell ) и Кэролин Кинг ( Carolyn M. King ). В их работах рассматривался вопрос, почему у горностая ( Mustela erminea ) эмбриональная диапауза наблюдается, а у малой ласки ( Mustela nivalis ) — нет. Тщательный обобщающий анализ был сделан в начале 2000-х годов Майклом Томом ( Michael Thom ) и другими учёными на основе построения филогенетического дерева куньих :112 .
Покой семян у растений
Эмбриональная диапауза у растений называется покоем семян . Под ним понимается состояние жизнеспособных семян , при котором они не прорастают. Если причиной покоя являются отсутствие условий для прорастания, такое состояние называется вынужденным покоем. Покой, связанный с особыми свойствами зародышей, называется органическим и проявляется в отсутствии прорастания или снижения всхожести при благоприятных для прорастания внешних условиях.
Существует три основных типа органического покоя семян :
- Морфологический — семена выпадают незрелыми, и перед прорастанием требуется некоторый период роста или дифференцировки .
- Физический — семена имеют водонепроницаемую оболочку, и для прорастания требуется нарушение её целостности.
- Физиологический — покой сохраняется до того момента, как в зародыше произойдут химические изменения.
Типы покоя могут совмещаться в одном семени. Широко распространён морфофизиологический покой — то есть одновременно относящийся и к морфологическому, и к физиологическому типу. Сочетание физического и физиологического покоя встречается редко, а сочетание морфологического типа с физическим невозможно.
Функциями покоя семян являются предотвращение всхода семян в условиях, которые благоприятны для всхода, но неблагоприятны для последующего роста и выживания растения, а также увеличение разнообразия вариантов развития — распространение семян в более широких ареалах и разновременный всход семян.
У некоторых растений семена в состоянии покоя могут находиться очень продолжительное время, вплоть до тысяч лет. Так, в 1951 году на северо-востоке Китая были обнаружены семена лотоса примерно тысячелетнего возраста, которые оказались жизнеспособны .
Эмбриональная диапауза у членистоногих
Среди беспозвоночных эмбриональная диапауза обнаружена у различных видов насекомых , ракообразных , и паукообразных ( клещей ) .
Диапауза (как эмбриональная, так и имажинальная ) широко распространена среди членистоногих и изучена у веслоногих ( дафний и артемий ); , и различных насекомых . Несмотря на то, что диапауза у членистоногих широко изучается в течение более полувека, на 2010 механизмы её регуляции до конца не поняты .
Эмбриональная диапауза у насекомых
У насекомых эмбриональная диапауза может происходить на различных стадиях развития эмбриона. Часто она называется «яйцевой диапаузой» ( англ. egg diapause , ovarian diapause ).
- Раннеэмбриональная (на ранних стадиях развития эмбриона).
- Среднеэмбриональная (на поздних стадиях развития эмбриона, после формирования нейроэндокринной системы ).
- Позднеэмбриональная (полностью развившаяся личинка, до выхода из яйцевой оболочки).
Однако у большинства видов диапауза происходит только на одной из указанных стадий.
В регионах с большими перепадами температуры диапауза позволяет пережить морозные зимы, либо периоды засухи за счёт большей устойчивости находящегося в состоянии диапаузы эмбриона к неблагоприятным условиям по сравнению со взрослыми особями.
Механизм диапаузы позволяет синхронизировать вылупление взрослых особей, увеличивая шансы на спаривание. Такая синхронизация важна, например, для долгоживущих видов наподобие , у которых личиночная стадия продолжается два лета, при этом скорость развития отдельных особей существенно различается.
Во многих случаях течение диапаузы определяется фотопериодом , испытанным материнским организмом. У некоторых видов, к примеру, у тутового шелкопряда ( Bombyx mori ) или ( Orgyia antiqua ) эмбриональная диапауза задаётся эмбриону «гормоном диапаузы», который вырабатывается в подглоточном ганглии матери и передаётся яйцу.
Наиболее подробно механизм эмбриональной диапаузы у насекомых был изучен у тутового шелкопряда ( Bombyx mori ). Вероятно, именно на этом организме лучше всего изучены механизмы подавления диапаузы. У ряда видов тутового шелкопряда наблюдается облигатная диапауза, такие виды являются . У других наблюдается факультативная диапауза, и они могут давать несколько поколений в течение одного года. Будет ли эмбрион такого вида проходить через стадию диапаузы, зависит от фотопериода, в течение которого развивался эмбрион его матери. Если эмбриональное развитие матери проходило в период высоких температур и большой продолжительности дня, то откладываемые ею яйца будут проходить через диапаузу на ранних стадиях развития. Если мать развивалась в период низких температур и коротких дней, её яйца будут развиваться без диапаузы. Таким образом, родившиеся весной, когда дни коротки, дадут потомство, выросшее летом и без диапаузы. Оно, в свою очередь, отложит яйца, которые пройдут через диапаузу и таким образом переживут зиму, чтобы развиться весной. В результате, такие виды оказываются бивольтинными, дающими два поколения в год.
Эндокринная связь между фотопериодом при развитии матери и диапаузой её яиц была выявлена Фукудой и Хасегавой в 1951 году . Гистологическими методами они показали, что если развитие зародышей и ранних личинок тутового шелкопряда при сравнительно высокой температуре (25 °C) и длинном световом дне, то в подглоточном ганглии развившихся из них куколок может быть выделен особый гормон диапаузы (обозначаемый DH ). Он приводит к увеличению содержания в их яйцах гликогена и пигмента ; такие яйца в своём развитии проходят через стадию диапаузы. Введение этого гормона женским куколкам приводит к тому, что они откладывают как диапазирующие, так и недиапазирующие яйца вне зависимости от того, какие яйца должны были быть отложены по условиям собственного развития.
В 1991 году гормон диапаузы тутового шелкопряда был выделен и синтезирован Имаи. Гормон имеет молекулярную массу 2645 а. е. м. , состоит из 24 аминокислот (TDMKDESDRGAHSERGALWFGPRL-амид), с амидированием на карбоксильном конце.
Однако маловероятно, что регуляция эмбрионального развития посредством гормона диапаузы является универсальным механизмом среди насекомых. Этот пептид был найден только у некоторых представителей чешуекрылых ( Lepidoptera ) .
Эмбриональная диапауза у ракообразных
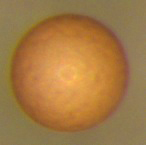
Одним из предметов изучения является рачок . Его диапазирующий эмбрион, также называемый цистой , способен выживать в экстремальных условиях . При комнатной температуре его диапазирующие эмбрионы могут обходиться без кислорода годами :306 .
На наступление и окончание эмбриональной диапаузы у ракообразных оказывают влияние различные факторы. Однако основным из них является — сезонные изменения продолжительности светового дня (например, для ветвистоусых раков ( Cladocera ), для ). Существуют и другие факторы, но обычно они действуют совместно с фотопериодом. :272-273 На выход из эмбриональной диапаузы влияют также различные факторы — изменения температуры, фотопериода и концентрации кислорода. Эти факторы действуют как на продолжительность летней, так и зимней диапаузы, но имеют различную степень важности. Так, фотопериод более важен для окончания летней диапаузы, а изменение температуры — зимней. :278
Эмбриональная диапауза способствует распространению многих ракообразных с балластными водами при судоходстве. И, возможно, именно эта физиологическая особенность позволила некоторым исходно морским видам раков заселить континентальные водоемы :58 .
У тех десятиногих ракообразных, у которых наблюдалась эмбриональная диапауза, она происходила в фазе гаструлы :43 .
У лат. Corystes cassivelaunus и Hyas coarcticus , имеющих одни из самых длительных периоды инкубации среди крабов - в 10 месяцев - эмбриональная диапауза обнаружена, и предполагается, что она возникла для увеличения выживаемости личинок, обеспечивая их развитие в периоды, когда пища максимально доступна :759 .
Эмбриональная диапауза у позвоночных
По всей видимости, у ряда различных представителей позвоночных эмбриональная диапауза возникла независимо в ходе эволюции в результате того, что виды, развитие зародышей которых могло приостанавливаться в зависимости от внешних условий, получили селективное преимущество :
- во-первых, диапауза позволяет задержать развитие эмбриона до того момента, пока внешние условия не станут более благоприятными для его выживания;
- во-вторых, диапауза позволяет увеличить промежуток времени между оплодотворением и рождением без изменения скорости процессов активного развития эмбриона.
Наблюдается среди карпозубых , черепах , сумчатых и высших зверей .
Яркие примеры диапаузы обнаружены у ряда карпозубых Африки и Южной Америки: представители родов , Aphyosemion , , Nothobranchius живут в водоёмах, которые существуют только в сезон дождей и полностью высыхают в остальное время. Взрослые особи мечут икру в сезон дождей, и эмбрионы погружаются в ил на глубине водоёма. С его высыханием все взрослые особи умирают, но клетки эмбрионов перед полным обезвоживанием частично отделяются друг от друга, рассеиваются и впадают в длительную диапаузу. Эта подготовка позволяет эмбрионам избежать полного иссушения посредством механизма, который пока ещё не изучен. Когда водоём в сезон дождей наполняется водой, смачивание эмбрионов водой восстанавливает их целостность, и их развитие возобновляется :250 .
Другая форма диапаузы наблюдается у некоторых видов морских черепах, у которых спаривание и оплодотворени может происходить за 3000 км до мест гнездовья. Развитие зародыша останавливается на стадии нейрулы на период миграции. В этой ситуации самка несёт в себе зародыш, не тратя энергию на поддержку его развития. Как только самка откладывает такие зародыши в виде «яиц», они продолжают свою программу развития в гнезде . Для части черепах характерно два периода эмбриональной диапаузы. Первый, происходящий до откладывания яиц, также называют "первичной диапаузой" или "расширением предяйцекладной задержки" ( англ. extension of preovipositional arrest ); он широко распространён среди черепах. Второй, наблюдаемый после откладывания яиц, называется собственно эмибриональной диапаузой .
У многих видов диапауза выражается только в уменьшении митотической активности на ранних стадиях развития эмбриона, а на последующие процессы развития влияния почти не оказывается. У подавляющего большинства млекопитающих скорость развития эмбриона не зависит от температуры, у остальных позвоночных и некоторых рукокрылых — зависит.
Эмбриональная диапауза у млекопитающих
У подавляющего большинства млекопитающих задержка развития эмбриона происходит только перед имплантацией в стенку матки . Для плацентарных эта задержка традиционно часто называется «задержанной имплантацией» , для сумчатых используется название «эмбриональная диапауза», так как возобновившая своё развитие бластоциста у них имплантируется в матку не сразу, в отличие от плацентарных .
Также у многих видов рукокрылых наблюдается задержка в развитии эмбриона после имплантации. Она называется «задержанное развитие» ( англ. delayed development ), обнаружена и среди представителей летучих мышей , и среди представителей крыланов . У ( Cynopterus sphinx ) этот процесс регулируется мелатонином .
Различают обязательную ( облигатную , сезонную ) и факультативную ( необязательную , лактационную ) эмбриональную диапаузу. В первом случае диапауза происходит в ходе каждой беременности (в частности, в определённое время года), а во втором случае — в условиях метаболического стресса (преимущественно при лактации ).
Из сумчатых, эмбриональная диапауза наблюдается у всех кенгуру , валлаби , и кенгуровых крыс , за исключением западного серого кенгуру ( Macropus fuliginosus ), кенгуру Лумхольтца ( Dendrolagus lumholtzi ) и кенгуровой мускусной крысы ( Hypsiprymnodon moschatus ), а также встречается у некоторых видов карликовых кускусов , карликовых летучих кускусов , и поссумов-медоедов . Даже у хищных сумчатых , у которых диапауза как таковая не наблюдается, в процессе развития бластоцисты происходит существенное изменение. Механизм регуляции диапаузы подробно изучен только для ( Macropus eugenii ).
Некоторым видам присущи оба механизма диапаузы — и обязательная, и факультативная, и они проявляются в зависимости от времени года. К таким видам относятся, например, таммары; механизм регуляции разных видов диапаузы различен: двустороннее удаление шейного ганглия у самок таммара устраняет сезонную эмбриональную диапаузу, но не оказывает влияния на лактационную.
Задержанная имплантация у млекопитающих
| Филогенетическое дерево для куньих. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
Обозначения:
|
Задержанная имплантация — это задержка развития эмбриона ряда плацентарных на стадии бластоцисты перед имплантацией в стенку матки. Корректность термина «задержанная имплантация» была поставлена под сомнение, но он продолжает использоваться наряду с предложенным для его замены термином «эмбриональная диапауза» .
Задержанная имплантация происходит не из-за случайного неприкрепления зародыша к слизистой оболочке матки, а из-за ингибирующего влияния матки на развитие бластоцист. Развивающееся яйцо, пройдя в яйцеводах период дробления и достигнув стадии бластоцисты, попадает в полость матки. Здесь оно некоторое время находится во взвешенном положении в жидкости просвета матки, без контакта с эпителием матки. У разных млекопитающих такая ситуация может длиться от нескольких часов (у хомяков ) и 2—3 дней (у человека) до нескольких месяцев (у соболя , броненосца и других) — последний упомянутый случай сопровождается эмбриональной диапаузой. Для начала процесса имплантации в матке должны произойти как общие изменения, определяющиеся гормональным фоном, так и локальные, связанные со взаимодействием с бластоцистой. В районе контакта с эмбрионом (или небольшого расстояния до него) клетки соединительно-тканной стромы матки преобразуются в децидуальные клетки . Вероятно, наряду с другими факторами, решающую роль в данном процессе играют иммунологические факторы.
Морфология остановившегося в развитии эмбриона различна у разных видов из числа тех, у которых происходит задержанная имплантация. У многих из них эмбрион перед диапаузой проходит процесс , при этом количество клеток в нём может составлять от 30—40 у косуль и 130 у мышей до 700 у девятипоясных броненосцев.
Задержанная имплантация была обнаружена у около сотни видов млекопитающих, относящихся к 7 отрядам, в 17 из 139 семейств : у парнокопытных ( Artiodactyla ), сумчатых ( Marsupalia ), насекомоядных ( Insectivora ), рукокрылых ( Chiroptera ), неполнозубых ( Edentata ), хищных ( Carnivora ) и грызунов ( Rodentia ). (В ряде источников со ссылкой на Айткена указывается цифра 8 отрядов; это связано с тем, что ранее ластоногие ( pinnipeds ) относились к отдельному отряду.) Облигатная диапауза выявлена по меньшей мере у 53 видов в 7 отрядах, 10 семействах ; при этом почти половина из этих видов относится к одному семейству — куньих . Задержанная имплантация наблюдается у 34 % куньих, в то время как в целом по млекопитающим она выявлена только для менее чем 0,05 %, что делает куньих предметом особого интереса исследователей.
Распространение задержанной имплантации среди млекопитающих носит нерегулярный характер. Значительное число подсемейств и даже родов включают как виды, у которых она наблюдается, так и виды, у которых её нет. Яркими примерами таких пар родственных видов, часто имеющих сходную среду обитания, но различающихся наличием и отсутствием задержанной имплантации, являются: канадская выдра ( Lontra canadensis ) и выдра обыкновенная ( Lutra lutra ); горностай ( Mustela erminea ) и ласка ( Mustela nivalis ); ( Spilogale gracilis ) и ( Spilogale putorius ) пятнистые скунсы (в каждой паре сначала идёт вид, у которого эмбриональная диапауза есть).
Существует подтверждённая исследованиями гипотеза, что задержанная имплантация преимущественно встречается в климате с выраженными сезонами, а также у долгоживущих организмов . Задержанная имплантация позволяет разнести процессы спаривания и вынашивания, фактически увеличивая период беременности. Это даёт возможность производить оба процесса в наиболее благоприятные для данного вида моменты, что повышает выживаемость потомства .
Гипотезы о возможности задержанной имплантации у человека
Несмотря на отсутствие прямых свидетельств того, что человеческий эмбрион может входить в состояние диапаузы, нельзя исключать возможность этого при определённых обстоятельствах.
Если удастся разработать метод искусственно вызывать диапаузу, это может увеличить:
- выживаемость эмбрионов после различных процедур;
- вероятность имплантации экстракорпорально оплодотворённого (ЭКО) эмбриона.
В нескольких исследованиях обнаружены задержки выработки хорионического гонадотропина при маточных и внематочных беременностях , хотя эти задержки скорее могут быть объяснены задержками в развитии, а не диапаузой.
В исследовании 1996 года описан случай задержки имплантации на 5 недель после ЭКО . Процесс проходил в условиях клиники с регулярным измерением уровня хорионического гонадотропина и проведением УЗИ . Эта пациентка ранее родила четырёх детей, причём основываясь на дате последней менструации, третий ребёнок родился на пять недель, а четвёртый — на две недели позже обычного срока человеческой беременности.
Исследования в этом направлении затруднены этическими соображениями. Исследования на макаках показали, что введение диклофенака (ингибитор синтеза простагландина ) вместе с (аналог простагландина Е 1 ) приводят к задержке имплантации до 6 суток, однако не было показано, что задержка вызвана именно диапаузой.
Механизм задержанной имплантации
В каждом семействе млекопитающих характеристики диапаузы различны, и регулирующие её факторы могут быть различными: диапауза может носить лактационный или сезонный характер, основываться на выработке эстрогена или прогестерона , зависеть от фотопериода или изменения состава пищи. Одним из основных гормонов, определяющих эндокринную картину у многих видов, является пролактин ; однако он может как стимулировать, так и ингибировать жёлтое тело .
|
← 4-OH-E 2 (ПГE 2 ) (цАМФ) |
|
|
EGFR ↑ ; COX-2 ↑ ; H 2 -R ↑ CB 1 ↓ |
EGFR ↓ ; COX-2 ↓ ; H 2 -R ↓ CB 1 ↑ |
|
|
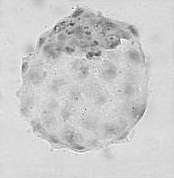
Молекулярные маркеры остановки и возобновления
развития бластоцисты грызунов:
|
||
Так, секретирование пролактина гипофизом предотвращает имплантацию у таммаров, но форсирует возобновление развития и имплантацию у норок и пятнистых скунсов. Удаление яичников (овариэктомия) — предотвращает диапаузу у куньих, но форсирует у армадиллов. Функция жёлтого тела остаётся постоянной у косуль и армадилло, но оно не завершает дифференциацию и выделяет низкий уровень прогестерона у хищных. Прогестерон вызывает имплантацию у таммаров, но у грызунов для этого требуется эстроген. У хищных же, по всей видимости, ни прогестерон, ни эстроген не может стимулировать имплантацию.
В регулировании сезонной диапаузы участвует мелатонин . Наиболее изучено его влияние для западных пятнистых скунсов, таммаров и норок, у которых удаление или денервация шишковидной железы приводит к отсутствию сезонной эмбриональной диапаузы, а введение мелатонина влияет на длительность диапаузы. Однако, по всей видимости, мелатонин не оказывает прямого воздействия на эмбрион, а вызывает изменение нейроэндокринной регуляции матери.
В период задержанной имплантации матка находится в состоянии покоя. Молекулярные механизмы, регулирующие процесс задержанной имплантации, изучены слабо.
На мышах и крысах проводили овариэктомию перед моментом выброса значительных количеств эстрогена , что приводило к развитию диапаузы бластоцисты в просвете матки. У грызунов состояние задержанной имплантации можно поддерживать продолжительное время при помощи прогестерона . При этом единичная инъекция эстрогена приводит к стремительной активации бластоцисты. Несмотря на то, что механизм регуляции этого процесса эстрогеном изучены мало, выявлены некоторые молекулярные маркеры .
Несмотря на наличие морфологических различий между бластоцистой в нормальном состоянии и в состоянии диапаузы, некоторые из молекулярных маркеров обратимо регулируют её стадии развития . Так, 4-гидроксиэстрадиол (4-OH-E 2 ), простагландин E 2 (ПГЕ 2 ) и цАМФ могут активировать покоящуюся бластоцисту in vitro и активировать, либо подавлять проявление маркеров, как указанно на иллюстрации. Если нормальная бластоциста переходит в состояние диапаузы перед прикреплением в матке, аналогичный процесс происходит в обратном направлении .
Список видов с выявленной эмбриональной диапаузой
| Вид |
Длительность
диапаузы |
Источник |
|---|---|---|
| Ракообразные | ||
| Десятиногие ракообразные | ||
| Большой сухопутный краб | 8 недель | :751 |
| Corystes cassivelaunus | 14 недель | :751 |
| Hyas coarcticus | 16 недель | :751 |
| Lopholithodes foraminatus | 12 месяцев | :13 |
| Maia squinado | 6 недель | :751 |
Комментарии
- В работе Bischoff, T. L. W. (неопр.) . — Rickersche Buchhandlung Gießen, 1854.
- Основываясь на следующей его работе: Ziegler, L. Beobachtungen über die Brunst und den Embryo der Rehe.. — Hannover: Hellweg`sche Hofbuchhandlung, 1843.
- ↑ Систематика периодически меняется. Указываемые числа — ориентировочные.
Источники информации
- ↑ John C. Avise. (неопр.) . — Cambridge University Press 2006. — P. 110. — 298 p. — ISBN 0521674174 .
- ↑ Flavia L. Lopes, Joëlle A. Desmarais and Bruce D. Murphy. (англ.) // Reproduction. — 2004. — Вып. 128 . — P. 669—678. 6 сентября 2008 года.
- Egg incubation: its effects on embryonic development in birds and reptiles. — Cambrige University Press.
- ↑ Mikael Sandell. (англ.) // The Quarterly Review of Biology. — Mar., 1990. — Т. 65 , № 1 . — P. 23—42.
- ↑ František Baluška. (неопр.) . — Springer, 2009. — P. 107. — ISBN 978-3-540-89229-8 .
- Диапауза / В. А. Свешников // Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров . — 3-е изд. — М. : Советская энциклопедия, 1969—1978.
- ↑ Michael D. Thom, Dominic D. P. Johnson and David W. Macdonald. (англ.) // Evolution. — Jan., 2004. — Т. 58 , № 1 . — P. 175—183. 14 августа 2011 года.
- ↑ М. B. Renfree, G. Shaw. (англ.) // Annual Review of Physiology. — Mar., 2000. — Т. 62 . — P. 353—375.
- ↑ Julian Lombardi. (неопр.) . — Springer, 1998. — P. 249. — ISBN 0792383362 , 9780792383369.
- ↑ Victor R. Alekseev, Jiang-Shiou Hwang, and Mei-Hui Tseng. (англ.) // Journal of Marine Science and Technology. — 2006. — Т. 14 , № 4 . — P. 269—286. (недоступная ссылка)
- ↑ G. Feldhamer, L. Drickamer, S. Vesery. (неопр.) . — The Johns Hopkins University Press, 2007. — P. 220. — ISBN 0801886953 .
- ↑ R. J. Aitken. (англ.) // Development in mammals. — 1977. — Т. 1 . — P. 307-308.
- ↑ Hermes, Robert. (неопр.) . — J. — Berlin: Diss. med.vet. FU-Berlin, 1997. 6 марта 2016 года.
- ↑ В. А. Заславский. . — Л. : Наука Ленингр. отд-ние, 1984.
- A. D. Lees. (неопр.) . — Cambrige: University Press, 1955.
- ↑ David Stanley Saunders. (неопр.) . — Oxford - New York: Pergamon Press, 1982. — P. 284.
- ↑ D. R. Khanna. (неопр.) . — New Delhi, India: Discovery Publishing House, 2004. — P. 291-315. — ISBN 817141897X , 9788171418978.
- ↑ Kinsaku Hasegawa. (англ.) // Nature. — 22 Jun., 1957. — Вып. 179 . — P. 1300—1301. 4 декабря 2016 года.
- Toshio Ichikawa, Satoshi Kamimoto. (англ.) // Zoological Science. — 2003. — Вып. 20 . — P. 979—983.
- ↑ H. Frederik Nijhout. (неопр.) . — Princeton University Press, 1998. — P. 166. — ISBN 0691059128 , 9780691059129.
- ↑ Julie A. Reynolds, Steven C. Hand. (англ.) // The Journal of Experimental Biology. — 12 June 2009. — Вып. 212(13) . — P. 2075-2084. — doi : . 10 января 2022 года.
- Tyndale-Biscoe, C. H. The role of the corpus luteum in the delayed implantation of marsupials (англ.) // Delayed Implantation / Enders, А. С. — University of Chicago Press, 1963. — P. 15—28.
- ↑ Patricia J. Berger. (англ.) // Nature. — 23 Jul., 1966. — Вып. 211 . — P. 435—436. 3 октября 2012 года.
- Carolyn M. King. (англ.) // Oikos 42. — 1984. — Вып. 20 . — P. 126—128. 24 мая 2010 года.
- М. Г. Николаева, М. В. Разумова, В. Н. Гладкова. . — Л. : Изд-во «Наука», 1985. 26 апреля 2010 года.
- ↑ Michael Fenner, Ken Thompson. (неопр.) . — Chicago Natural History Museum, 2005. — P. 97.
- (неопр.) . — Springer, 1953.
- ↑ Takashi Homma, Ken Watanabe, Shuichiro Tsurumaru, Hiroshi Kataoka, Kunio Imai, Mari Kamba, Teruyuki Niimi, Okitsugu Yamashita and Toshinobu Yaginuma. G protein-coupled receptor for diapause hormone, an inducer of Bombyx embryonic diapause (англ.) // Biochemical and Biophysical Research Communications. — San Diego, CA, USA: Elsevier, May 26, 2006. — Т. 344 , вып. 1 . — ISSN .
- Richard W. Hill, Gordon A. Wyse, and Margaret Anderson. (неопр.) . — Sinauer Associates, Inc, April 4, 2008. — ISBN 978-0-87893-317-4 . (недоступная ссылка)
- Julie Annette Reynolds. [ Animal Metabolism and Gene Expression During Embryonic Diapause in Arthropods (A Dissertaion)] (неопр.) . — December 2007. 2 июля 2010 года. . Дата обращения: 27 декабря 2010. Архивировано 2 июля 2010 года. .
- ↑ R. F. Chapman. (неопр.) . — Cambridge University Press, 1998. — ISBN 0521578906 .
- (недоступная ссылка) на сайте «Эмбриология животных», 18.10.2009
- Kunio Imai, Takamichi Konno, Yoshitaka Nakazawa, Takashi Komiya, Minoru Isobe, Kazushi Koga, Toshio Goto, Toshinobu Yaginuma, Kiyoshi Sakakibara, Kinsaku Hasegawa, Okitsugu Yamashita. (англ.) // Proceedings of the Japan Academy. Ser. B: Physical and Biological Sciences. — 1991. — Т. 67 , № 6 . — P. 98-101. — ISSN . — doi : . (недоступная ссылка)
- Robert A. Browne, Patrick Sorgeloos, Clive N. A. Trotman. (неопр.) . — CRC Press, 1991. — P. 94-118. — 374 p. — ISBN 0849367298 , 9780849367298.
- Menze M. A., Fortner G., Nag S., and Hand S. C. (англ.) // Apoptosis. — 2010. — Т. Mar;15(3) . — P. 293-312. 9 апреля 2012 года.
- Vadim E. Panov, Piotr I. Krylov, and Nicoletta Riccardi. (англ.) // Journal of Limnology. — 2004. — Т. 63 (Suppl. 1) . — P. 56—69. (недоступная ссылка)
- ↑ William Duguid. Laboratory study of reproduction and development of Lopholithodes foraminatus (brown box crab), with a discussion of reversed asymmetry. — University of Victoria, 2002.
- ↑ Robert G. Wear. Incubation in British Decapod Crustacea, and the Effects of Temperature on the Rate and Success of Embryonic Development (англ.) // Journal of the Marine Biological Association of the United Kingdom. — 1974. — Vol. 54 . — P. 745-762 .
- David T. Booth. (англ.) // Journal of Herpetology. — Society for the Study of Amphibians and Reptiles. — Т. 36 , № 2 .
- ↑ Rodney A. Mead. (англ.) . — 1993. — Т. 266 , № 6 . — P. 629—641. (недоступная ссылка)
- Arnab Banerjee, K. J. Meenakumari, S. Udin and A. Krishna. (англ.) // Reproduction. — 2009. — Вып. 138 . — P. 935—944. (недоступная ссылка)
- C. Hugh Tyndale-Biscoe. (неопр.) . — Victoria, Australia: CSIRO Publishing, 2005. — 442 p. — ISBN 0-643-06257-2 .
- Marilyn B. Renfree, D. W. Lincoln, O. F. X. Almeida, R. V. Short. (англ.) // Nature. — 10 Sep., 1981. — Вып. 293 . — P. 138—139.
- И. Н. Вагина, С. В. Евсиков, А. П. Соломко. // Біополімери і клітина. — 2003. — Т. 19 , № 1 . — ISSN . 4 марта 2016 года.
- Б. П. Токин. Общая эмбриология: Учеб. для биол. спец. ун-тов. — 4-е изд., перераб. и доп. — М. : Высшая школа, 1987.
- ↑ Jørgen Grinsted, Birthe Avery. (англ.) // Human Reproduction. — 1996. — Т. 11 , № 3 . — P. 651–654.
- ↑ Juan J. Tarín, Antonio Cano. (англ.) // Human Reproduction. — 1999. — Т. 14 , № 10 . — P. 2434–2436.
- James Olcese. (неопр.) . — Springer, 2000. — P. 206. — 488 p. — ISBN 030646134X , 978-0306461347.
- ↑ Ernst Knobil, Jimmy D. Neill. (неопр.) . — Elsevier Academic Press, 2006. — P. 154.
Литература
- John C. Avise. (неопр.) . — Cambridge University Press 2006. — P. 110-113. — 298 p. — ISBN 0521674174 .
- Flavia L. Lopes, Joëlle A. Desmarais and Bruce D. Murphy. (англ.) // Reproduction. — 2004. — Вып. 128 . — P. 669—678. 6 сентября 2008 года.
Ссылки
- - The Diapause Research Foundation, некоммерческая организация, занимающаяся изучением механизмов диапаузы и поддержкой других квалифицированных учёных, работающих в этой области.
| Стадии |
|
|---|---|
| Процессы | |
| Зародышевые листки | |
| Дифференцировка клеток | |
- 2020-12-31
- 1

