Хорошие статьи
- 1 year ago
- 0
- 0


Фитотельма́ты (от др.-греч. φῠτόν — «растение» + τέλμα, ατός — «стоячая вода», «болото») — экосистемы небольших водоёмов, образованные в листьях и цветках растений или дуплах деревьев . Название этому типу экосистем дал в 1928 году немецкий биолог Людвиг Варга.
Фитотельматы можно встретить в различных природных зонах от экваториальных лесов до Субарктики . Фитотельматы образуются более чем 1500 различными видами растений из 26 семейств. В зависимости от систематической принадлежности и органов растений, в которых они формируются, выделяют несколько групп фитотельматов. Видовой состав обитателей этих экосистем зависит от различных факторов: объёма воды, уровня освещения, концентрации питательных веществ (главным образом азота ) и др. Во многих группах живых организмов, населяющих фитотельматы, имеются виды, не встречающиеся в других типах водных экосистем . По палеонтологическим данным, экосистемы фитотельматов образовывались уже в эоцене , о чём говорит обнаружение водных организмов в янтаре .
Термин «фитотельматы» предложил немецкий биолог Людвиг Варга (Ludwig Varga) в 1928 году в статье «Ein interessanter Biotop der Biocönose von Wasserorganismen» («Интересный биотоп биоценозов водных организмов»), вышедшей в журнале «Biologisches Zentralblatt». Активное использование этого термина в англоязычной литературе началось после выхода в 1971 году статьи американского зоолога . Варга детально изучил видовой состав растений и животных, обитающих в пазухах листьев ворсянки , а также обобщил известные на тот момент факты о заселении подобных местообитаний. Так, уже было известно, что разнообразные организмы заселяют листья в форме кувшина у некоторых видов хищных растений, обитающих в тропической Азии ( непентес ) и Северной Америки ( саррацения ). К фитотельматам Варга отнёс и заполненные водой пазухи веток бука . До работ Варги учёные почти не обращали внимание на этот класс местообитаний, хотя о развитии кровососущих комаров в листьях растений упоминалось ещё в китайских трактатах династии Тан (между 618 и 905 годами н. э.). В конце XIX века о развитии водных животных на деревьях писал Фриц Мюллер . В 1904 году австралийский натуралист привёл первые сведения об обитателях листьев цефалота . В 1922 году русский энтомолог Владимир Алпатов изучал фауну микроводоёмов во влагалищах листьев дудника лесного . На работу Варги обратил внимание Август Тинеманн и в 1932 году изучил фауну листьев-кувшинок непентеса .
Фитотельматы формируются у более чем 1500 различных видов растений из 26 семейств . К началу XXI века установлено, что фитотельматы характеризуются довольно высоким разнообразием групп живых организмов. Однако отмечается, что в действительности все виды никогда не встречаются одновременно в отдельной единице среды обитания. Численность разных видов может быть различна, некоторые редкие виды могут быть случайными обитателями. Таким образом, пищевая сеть, образованная только доминирующими видами, оказывается намного проще, чем та, в которой учитываются все виды без учёта их обилия. Пищевые предпочтения многих видов, отмеченных в фитотельматах, пока не известны .
Фитотельматы встречаются во многих природных зонах от экваториальных лесов до Субарктики . Австралийский биолог в 1987 году разделил фитотельматы по способу формирования на пять основных групп: 1) полости или углубления деревьев (дендротельматы); 2) междоузлия бамбука; 3) водоёмы в листьях, прицветниках или лепестках растений; 4) растения-кувшины; 5) резервуары бромелиевых .
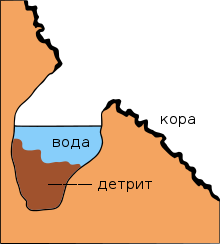
Дендротельматами называют заполненные водой полости внутри дерева или углубления на его поверхности. Они наиболее широко представлены в тропических широтах. Выделяют две формы дендротельматов: «кюветы» и «дупла». В первом случае углубления образуются при физической деформации дерева под воздействием внешних факторов, без повреждения коры. Во втором варианте отверстия образуются за счёт развития гнилостных процессов внутри дерева. Объём дендротельматов может быть от нескольких миллилитров до 30 литров. Большие водоёмы могут сохраняться в течение длительного времени — например, резервуары в деревьях , произрастающих во влажных тропических лесах на востоке Австралии , могут существовать более 50 лет. Основу трофической цепи составляет детрит , образующийся на дне дендротельматов при разложении органических веществ. На видовой состав фауны влияют кислотность и температура воды, содержание в ней танинов и лигнинов . Пищевые цепочки в этих экосистемах короткие. Наиболее хорошо изучена фауна кровососущих комаров, развивающихся в дендротельматах. В микробном комплексе преобладают Proteobacteria и Bacteroidetes . Вещества, выделяемые бактериями, стимулируют развитие комаров рода Aedes , являющихся переносчиками лихорадки денге и жёлтой лихорадки . В дендротельматах клёна в Норвегии встречается 139 видов микроскопических грибов . В Южной Индии в дуплах деревьев развиваются личинки стрекозы Lyriothemis tricolor .

Образование фитотельматов в стеблях бамбука происходит, если стебель сломан в результате естественных причин или срублен человеком. Фитотельматы могут образоваться и в междоузлиях живых растений бамбука. Жидкость, представляющая выделения растения, смешанные с дождевой водой, затекает туда через небольшие отверстия диаметром около 1 см, которые проделывают жуки-листоеды и усачи . Отсеки междоузлий водонепроницаемы и это способствует длительному сохранению воды в этих резервуарах. Объём таких фитотельматов может достигать 1 литра. Сообщества организмов могут формировать здесь довольно сложные пищевые цепи, длина которых зависит от степени открытости фитотельмата и объёма питательных веществ, накопленного в нём .
Фитотельматы в листьях и прицветниках образуются преимущественно у однодольных растений из семейств коммелиновых , пальмовых и лилейных . Максимальный объём воды достигает 100 мл , поэтому пищевые цепи в этих биотопах очень короткие. Несмотря на небольшой объём, эти резервуары активно заселяются насекомыми. В Новой Гвинее все соцветия куркумы ( ) были заселены личинками кровососущих двукрылых. Прицветники геликонии , обитающей в Южной Америке , могут содержать до 20 мл воды. Эта вода содержит нектар и части цветков, которые разлагаются бактериями. Из животных наиболее часто в этих фитотельматах встречаются лёгочные моллюски , личинки двукрылых и жесткокрылых насекомых и некоторые лягушки ( Ranitomeya ventrimaculata ) . В прицветниках геликонии обитает специфическая инфузория . Обилие этой инфузории зависит от количества личинок комаров, которые ею питаются. Перенос инфузорий с одного растения на другое осуществляется улитками и двукрылыми. В умеренных широтах такие фитотельматы чаще образуются на двудольных , — например, дягиле и ворсянке . Кроме того, фитотельматы образуются в розетках листьев пандановых . На Гавайских островах в них развиваются два эндемичных вида стрекоз — и Megalagrion koelense .


Растения-кувшины имеют видоизменённые листья, которые способны собирать воду. Эти растения насекомоядны , они встречаются в Австралии ( Cephalotaceae ), Южной Азии ( Nepenthes ) и Северной Америке ( Sarraceniaceae ). Насекомые, тонущие в этих водоёмах, разлагаются или ферментами , выделяемыми самим растением, или обитающими в них бактериями. В кувшинах Cephalotus follicularis в Австралии обитают личинки беспозвоночных и одноклеточные водоросли рода . Из насекомых в кувшинах развиваются личинки эндемичного для юго-запада Австралии вида мух-ходуленожек Badisis ambulans . Личинки питаются разлагающейся добычей кувшина и окукливаются в нём. Задние дыхальца у личинки атрофированы , поэтому она поглощает кислород из жидкости кувшина всей поверхностью тела . Объём кувшина непентеса достигает двух литров. В фитотельматах встречаются семь видов раковинных амёб : , , , , , и . С возрастом в фитотельматах увеличивается объём органики, и это приводит к повышению видового разнообразия беспозвоночных, обитающих в них. Из растений-кувшинов Северной Америки только у Sarracenia purpurea могут формироваться устойчивые длительно существующие сообщества, поскольку другие виды обитают в засушливом климате и вода в кувшинах быстро испаряется . В фитотельматах непентесов формируются довольно сложные взаимоотношения между их обитателями. Основной ресурс, за который идёт борьба между обитателями фитотельматов, — это растворимый азот . Было показано, что в фитотельматах, в которых имеются хищники, концентрация азота выше, чем в тех, откуда они были удалены .

Бромелиевые , многие из которых являются эпифитами , встречаются преимущественно в Центральной и Южной Америке. Фитотельматы расположены в основании розеток листьев и существуют круглый год, позволяя развиваться многочисленным, специфичным и разнообразным в видовом отношении водным организмам . В Коста-Рике в фитотельматах бромелий уже к 1913 году было замечено около 400 видов беспозвоночных . Фитотельматы бромелиевых служат местом развития яиц и головастиков лягушек древолазов (семейство Dendrobatidae ). Через некоторое время после откладки икры́ самка лягушки возвращается к тому же фитотельмату и откладывает в него неоплодотворённые икринки, которые служат пищей развивающимся головастикам. Часто в одном резервуаре до взрослой стадии доживает только одна особь из-за выраженного каннибализма , характерного для головастиков этих лягушек . Разнообразие беспозвоночных, обитающих в фитотельматах, зависит от открытости отверстия и объёма фитотельмата, расстояния от земли и от температуры воды . Увеличение биоразнообразия с высотой учёные связывают с растущей интенсивностью солнечного света и большим накоплением дождевой воды. Более крупные бромелии обладают большей водоудерживающей способностью, что увеличивает время, необходимое для завершения цикла развития его обитателей .
Часто фитотельматы образуются также в упавших плодах, листьях и прицветниках растений, например, скорлупе кокосовых орехов , плодов какао . В воде, накапливающейся в этих резервуарах, обитают личинки беспозвоночных и головастики лягушек-древолазов . Крупные листья некоторых тропических деревьев при высыхании скручиваются и могут наполнятся водой. Примерами могут быть листья какао, кокоса , текстильного банана , диллении кустарниковидной и . На острове Майотта , около Мадагаскара , в расколотых кокосовых плодах развиваются кровососущие комары , Aedes aegypti ( комар жёлтолихорадочный ) . На Калимантане в упавших кокосовых плодах с отверстиями, проделанными крысами или тупайями , обитают личинки комаров рода Armigeres .
В сообществах фитотельматов и в Юго-Восточной Азии преобладают Proteobacteria ; кроме них, здесь регулярно отмечали Actinobacteria (15,8 %), Bacteroidetes (17,2 %) и Firmicutes (8,8 %) . Бактерии из семейства являются пищей для личинок комаров . Среди экологических групп бактерий значительную роль играют азотфиксирующие бактерии порядков ( , ) и ( ). В жидкости кувшина встречали представителей семейства Enterobacteriaceae и родов Lactobacillus , и , которые являются преимущественно кишечными симбионтами позвоночных и насекомых. На внешних стенках кувшина идентифицированы бактерии рода Clostridium . Численно доминирующей группой в жидкости были ацидофильные бактерии рода , составлявшие до 74 % от общего числа клеток микроорганизмов. Это объясняется очень высокой кислотностью среды обитания — от 2,1 до 2,7 pH . У представителей родов Aechmea и (семейство Бромелиевые ) в Коста-Рике состав бактериального сообщества фитотельматов зависит от кислотности среды. В резервуарах с pH жидкости выше 5,3 преобладали , Firmicutes и Bacteroidetes . В фитотельматах с pH менее 5,1 преобладают , Acidobacteria , Planctomycetes и Bacteroidetes .
В фитотельматах мексиканских бромелий Bromelia и Tillandsia обнаружено 27 видов инфузорий , из которых были наиболее широко распространены виды , и Tetrahymena sp. Большинство видов, обнаруженных в фитотельматах, встречаются также в почве и обычно являются космополитами , а вид , найденный только в фитотельматах, ограничен в распространении неотропической областью . В Бразилии в фитотельматах обнаружены 92 вида инфузорий, принадлежащих 14 отрядам . Видовое разнообразие инфузорий в засушливый сезон было выше, чем в сезон дождей .
Среди микроскопических грибов в фитотельматах обнаружены представители гифомицетов и . Гифомицеты участвуют в разрушении органического вещества и улучшении питательных свойств опада для обитающих в фитотельматах беспозвоночных и головастиков земноводных. Грибные колонии дендротельматов могут служить источником спор для водных грибов, которые колонизируют органическое вещество в ручьях . В дуплах клёна в Норвегии наиболее часто встречаются , , , , и виды рода Alternaria . Специфическими гифомицетами в заполненных водой дуплах бука , дуба и ясеня в Венгрии являются , и .
Большинство трихомицетов связаны с личиночными стадиями водных беспозвоночных. Между беспозвоночными и трихомицетами, как правило, формируются отношения типа комменсализм , мутуализм или паразитизм , при этом грибы семейства обитают в задней кишке, а представители семейства прикрепляются к , выстилающей среднюю кишку . В Южной Америке в фитотельматах бромелиевых и зонтичных наиболее часто встречаются и .
Грибы хитридиомицеты могут паразитировать на икре лягушек, развивающихся в фитотельматах. Повторно посещая кладки, лягушки выделяют в воду вещества, которые подавляют рост патогенных грибов .
В фитотельматах бромелии , растущей в хорошо освещённых местообитаниях во Французской Гвиане , численность водорослей варьировала от 9,5×10 3 до 2,6×10 5 клеток/мл. Биомасса их составляла около 30 % от общей микробной биомассы сообщества. Отмечается, что водоросли составляют основу рациона питания живущих в фитотельмате личинок кровососущих комаров . Видовой состав эвгленовых водорослей в фитотельматах бромелий , , и изучался в Бразилии. Относительно более щелочную среду (pH около 6) предпочитает , хотя она была обнаружена и в фитотельматах с более кислой средой. Наиболее ацидофильным видом эвгленовых был , развивающийся при pH около 3,3. Помимо кислотности на состав и обилие эвглен влияют освещённость и наличие органики . При затенении многие виды этой группы становятся бесцветными и переходят на сапротрофное питание .
В фитотельматах обитает большое число групп беспозвоночных, в том числе коловратки , ракушковые и веслоногие ракообразные , плоские черви , нематоды , олигохеты , пиявки , насекомые и паукообразные , многие из которых не определены до вида . Фитотельматы, в которых могут обитать беспозвоночные, формируются у цветковых растений из 31 семейства. Многие группы беспозвоночных, отмеченные в фитотельматах, с точностью до вида не определены .
В бромелиях в Коста-Рике и на Ямайке обнаружены плоские черви из семейств Geoplanidae ( ), ( и ) и ( и ). Эти черви встречаются обычно по краям фитотельмата, где выше уровень растворенного кислорода. Планарии рода в дуплах деревьев Австралии питаются личинками комаров . В бромелиевых в Аргентине и дуплах деревьев в Германии известны находки брюхоресничных червей рода . Они питаются, вероятно, мелкими частицами органического вещества .
Живущие в фитотельматах бромелий нематоды и из отряда рабдитид ( , , ) питаются бактериями. Представители родов , , потребляют микроскопические грибы. Беспозвоночных потребляют дориляймиды родов , , . Простейшими питаются эноплиды рода . Преобладающими во видовому разнообразию группами являются группы бактериофагов и хищников. Максимальная численность нематод отмечена в период с октября по апрель .
Во всех классах фитотельматов обычно встречаются олигохеты . В фитотельматах бамбука и дендротельматах 17 видов древесных растений на острове Цусима (Япония) были собраны кольчатые черви из семейства Naididae , но в целом в литературе отмечается, что черви не играют большой роли в сообществах фитотельматов . Многие малощетинковые черви участвуют в разложении органических веществ. Пиявки , обнаруженные в бромелиевых, вероятно, используют их как временное убежище. Единственный специализированный обитатель фитотельматов найден в пазухах листьев банана и алоказии крупнокорневищной в Юго-Восточной Азии и на Новой Гвинее . Предположительно, он питается личинками насекомых и олигохет .
В фитотельматах разных типов обитают более 100 видов ракообразных . Наибольшим видовым разнообразием и численностью обладает отряд Copepoda , представленный в фитотельматах 68 видами. Заметную роль играют также Decapoda (16 видов) и Ostracoda (14 видов). Самой небольшой группой являются Cladocera , из которых только 10 видов живут в фитотельматах. Ракушковые ракообразные обитают только в дендротельматах умеренного пояса и фитотельматах бромелиевых в Неотропической области. В переносе многих ракообразных между фитотельматами участвуют земноводные и некоторые другие виды животных. Для некоторых групп ракообразных, особенно для дафний , важным фактором распространения может быть ветер .
Впервые ракообразные в водоёмах в дуплах бука были отмечены в 1920 году, это виды и . С бромелиевыми связаны преимущественно представители рода , а также , , и и др. Некоторые представители Ostracoda, живущие в фитотельматах, морфологически сходны с видами, обитающими в подземных водах и в воде порах почвы. Веслоногие ракообразные обитают в фитотельматах всех типов. В рамках этой группы наибольшую связь с фитотельматами имеют представители Harpacticoida . Из 28 видов гарпактицид в фитотельматах 20 видов встречаются только в этом местообитании .
Шесть видов ветвистоусых относятся к семейству Chydoridae и являются типичными представителями бентоса , а четыре вида семейства Daphniidae обитают в толще воды. Виды , и отмечены в дуплах деревьев. Они, вероятно, являются, случайными обитателями фитотельматов, поскольку не имеют специальных приспособлений для жизни в этой среде, кроме того, они часто встречаются в крупных водоёмах. Остальные семь видов Cladocera отмечены только в фитотельматах бромелиевых . Типичным представителем облигатных обитателей фитотельматов является вид , описанный из Никарагуа . Он имеет сплющенные с боков усики и короткий базальный шипик на коготках у самок . Численность этого вида может достигать 45 особей на литр .
Большинство крабов найдены в фитотельматах в Азии и Вест-Индии . Они, как правило, слабо связаны с фитотельматами, но некоторые виды используют эти местообитания для размножения или в качестве укрытий. Краб , обитающий на Ямайке, поселяется в фитотельматах бромелии . Личинками крабов питаются личинки стрекозы . В течение суток они могут уничтожить до 5 личинок краба .
Пауки в фитотельматах встречаются редко. В Бразилии при обследовании фауны фитотельматов бромелий и обнаружен паук-птицеед . Над одним из листьев паук плетёт навес из паутины и остаётся за ним. В случае опасности паук скрывается в фитотельмате, иногда полностью погружаясь в воду. Таким образом, паук получает убежище, а бромелия использует остатки пищи паука как источник азота . В фитотельматах обитают пауки-бокоходы Thomisus nepenthiphilus и . Было показано, что пауки оказывают существенное влияние на динамику численности обитателей фитотельматов .

Насекомые являются наиболее изученной группой обитателей фитотельматов, в которой отмечены представители шести отрядов: стрекозы , ручейники , полужесткокрылые , двукрылые , жесткокрылые и перепончатокрылые .
В тропических регионах с фитотельматами связаны представители 31 рода стрекоз из четырёх семейств: (1 род), Aeshnidae (3 рода), Libellulidae (8 родов), Coenagrionidae (18 родов) . В пазухах листьев панданов на Новой Гвинее обитают 28 эндемичных видов стрекоз-стрелок рода Papuagrion . Их предполагаемые предки из рода обитают в стоячих водоёмах с плохо аэрируемой водой, что послужило предпосылкой к заселению ими фитотельматов . Приспособлением к недостатку кислорода являются расширенные хвостовые жабры . В кувшинах бромелиевых в Южной Америке живут представители рода Bromeliagrion и . В Панаме в дуплах деревьев встречаются стрекозы Gynacantha membranalis , , и . В Восточной Африке в дендротельматах развиваются стрекозы Coryphagrion grandis . В Африке в фитотельматах обнаружены ручейники из семейств Limnephilidae , Conoesucidae и Leptoceridae .
В листьях бромелиевых многочисленными являются клоп-гребляк и гладыш . Иногда встречаются представители семейств Pleidae и Belostomatidae . В Малайзии в бамбуковых пнях и междоузлиях найдены представители семейств Veliidae , Hydrometridae и Gerridae .
Жизнедеятельность муравья Colobopsis schmitzi тесно связана с кувшинами . Эти насекомые способны передвигаться по скользким краям кувшина, нырять в его жидкость и плавать в ней без вреда для себя. Они питаются насекомыми, попавшими внутрь фитотельмата, и потребляют нектар, выделяющийся на поверхности кувшина .
Большую роль в сообществах фитотельматов играют личинки двукрылых . Согласно наблюдениям в Юго-Восточной Азии и Австралии, в составе этих экосистем встречаются представители 20 семейств . Широко представлены в фитотельматах комары-звонцы . К наиболее распространённым родам звонцов в фитотельматах относятся , и Monopelopia , иногда встречаются Chironomus и . В жидкости обитают хищные личинки из семейства грибных комаров . На поверхности жидкости они выделяют липкую паутинную сеть для отлавливания имаго комаров Phoridae и Culicidae , также обитающих в фитотельматах. Грибные комары поселяются в непентесе после заселения его другими комарами, поэтому они предотвращают вылет из кувшина взрослых комаров и этим способствуют сохранению питательных веществ внутри фитотельмата .
В фитотельматах различных типов обитают личинки кровососущих комаров. По оценкам специалистов, только в фитотельматах бромелий встречаются более 200 видов этого семейства. Наиболее тесно связаны с фитотельматами представители родов Culex (подрод ), Aedes (подрод ), Wyeomyia и Anopheles (подрод ). Комары, развивающиеся в фитотельматах, часто являются переносчиками опасных заболеваний, например, лихорадки денге и филяриатозов .
Из жуков в фитотельматах встречаются представители семейств трясинников , водолюбов и плавунцов . Специализированными обитателями фитотельматов бромелиевых являются плавунцы родов Desmopachria и . Эти роды жуков появились одновременно с бромелиевыми приблизительно 12—23 млн лет назад .

В 1927 году американский зоолог описал откладывание яиц в фитотельматы эндемичной для Танзании лягушкой . Это первое свидетельство использования бесхвостыми амфибиями микроводоёмов в листьях растений для размножения. Поначалу этот факт воспринимался как отклонение от нормы. В дальнейшем выяснилось, что это довольно распространённое явление в тропических лесах. К 2004 году было отмечено уже более 100 подобных видов из 9 семейств и 44 родов .

На Тайване лягушки Kurixalus eiffingeri откладывают яйца в фитотельматы, образующиеся в пнях бамбука . Эти яйца поедаются хищным слизнем и поражаются хитридиомицетами. Самки лягушек неоднократно посещают кладку для откладки неоплодотворённых яиц, которыми питаются головастики. Кроме того, кладки посещаются самцами. Если препятствовать самцам посещать кладки, то увеличивается смертность яиц от паразитов и хищников. Было замечено, что самец распознаёт слизней как хищников и удаляет любые другие подозрительные объекты, которые могут повредить яйцам. Кожные выделения самца, попадающие в воду, ограничивают рост паразитических грибов .
Растение , растущее на острове Калимантан , практически не посещается насекомыми. Однако над жидкостью, скапливающейся в кувшинах этого растения, регулярно были замечены на ночёвке летучие мыши Kerivoula hardwickii . Взаимоотношение этих видов является примером мутуализма . Расстояние от поверхности жидкости до края кувшина составляло около 15 см , что представляет идеальное укрытие для этих животных. Растение получает азот из фекалий этих летучих мышей. Расчёты показывают, что непентес получает таким способом почти 38 % от необходимого ему азота .
В горных районах острова Калимантан встречается вид , который образует кувшины двух видов. Молодые растения имеют обычные плотоядные кувшины, добычей которых служат членистоногие . Кувшины зрелых растений не имеют приспособлений для питания членистоногими. Они выделяют экссудаты , которые накапливаются на крышке кувшина. Этими выделениями питаются мелкие древесные млекопитающие тупайи , экскременты которых во время питания падают в кувшин непентеса. Было рассчитано, что за счёт экскрементов тупай растение получает от 57 до 100 % необходимого ему азота .
Сведения о существовании фитотельматов в прошлые геологические эпохи очень скудны, однако обнаружение типично водных насекомых в ископаемых янтарях говорит о существовании полостей, заполненных водой, в деревьях, из смолы которых образовался янтарь . В доминиканском янтаре , датируемом верхним эоценом , обнаружены клоп и кровососущий комар .