Interested Article - Апторнис
- 2021-09-25
- 1
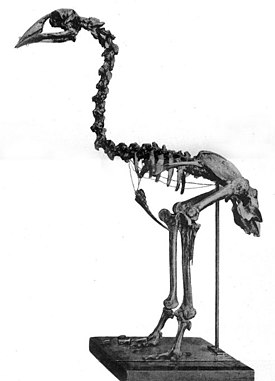
Апторнис ( лат. Aptornis ) — род вымерших птиц отряда журавлеобразных . Крупные наземные птицы высотой около 80 см и массой до 25 кг обитали на островах Новой Зеландии с раннего миоцена до XIII века.
Род был описан Гидеоном Мантеллом в 1848 году. В XIX веке он включал два вида: Aptornis defossor с Южного острова и Aptornis otidiformis с Северного острова, в XXI веке был также описан Aptornis proasciarostratus .
Обнаружение
Первый апторнис был описан английским палеонтологом Ричардом Оуэном в 1843 году на основании тибиотарзуса и бедренной кости , найденных на восточном побережье Северного острова . Оуэн предположил что находка является ещё одним видом моа и назвал её Dinornis otidiformis . С обнаружением большего числа ископаемых остатков, в том числе фрагментов плюсны и черепа , систематика была пересмотрена и вид выделен в отдельный род Aptornis .
В 1849 году в известковой пещере близ города на северо-западе Северного острова в 30 км от берега был обнаружен полный и почти полный скелеты апторнисов. К 1871 году на северном побережье Южного острова Оуэн обнаружил 105 костей, принадлежащих по меньшей мере восьми особям, и описал новый вид A. defossor , к которому отнёс почти полный скелет, найденный в 1849 году .
В 1888 году близ деревни на Южном острове был обнаружен череп птицы, похожей на апторниса, но большего размера. Оуэн дал этим остаткам название Aptornis bulleri , но не предоставил формального описания. Данный таксон остаётся nomen nudum , а некоторые учёные полагают, что разница в размерах связана с половым диморфизмом .
Более поздние находки апторнисов связаны с исследованиями австралийского палеозоолога . В 2011 году в регионе Южного острова Уорти с соавторами на основании двух грудных позвонков описал третий вид апторнисов — A. proasciarostratus . К этому виду были отнесены фрагменты бедровой кости, фаланги, позвонка и, возможно, тибиотарзуса, найденные в том же месте раскопок .
Описание
Апторнисы были высотой около 80 см . Более мелкие A. otidiformis имели массу до 16 кг , а более крупные A. defossor — около 19 кг ; некоторые учёные предполагают, что их масса могла достигать 25 кг . Самым маленьким апторнисом был A. proasciarostratus , обитающий обособленно на Южном острове .
Учёные считают, что апторнисы были наземными птицами, не способными к полёту . Они обладали очень маленькими крыльями, массивным черепом и клювом. Их крепкие шейные позвонки указывают на развитую мускулатуру, а толстые ноги и лапы скорее всего использовались вместе с клювом чтобы выкапывать или фиксировать добычу .
Помимо своих малых размеров A. proasciarostratus отличается от остальных апторнисов повышенной пневматичностью скелета и некоторыми характеристиками ( англ. processus spinosus ) и ( англ. processus transversus ) .
Распространение
Апторнисы обитали на Северном и Южном островах Новой Зеландии с раннего миоцена до XIII века. Птиц Южного острова учёные относят к виду Aptornis defossor , птиц Северного острова — Aptornis otidiformis . Описанный в 2011 году A. proasciarostratus обитал в районе округа Южного острова. Его остатки также датируются ранним миоценом (19—16 млн лет назад ) и являются самыми старыми из известных апторнисов .
По местам обнаружения остатков учёные предположили, что апторнисы обитали на побережье островов. В XIX веке A. defossor был обнаружен в Те-Куити на Северном острове и в многочисленных пещерах на холме на северном побережье Южного острова, а также в пещерах в 20-25 км от Оамару . Несмотря на большое количество хорошо сохранившихся остатков Оуэн посчитал апторнисов «редкими» ( англ. rare ). Спустя сотню лет Уорти уточнил, что ископаемые остатки апторнисов преимущественно относятся к голоцену , в то время как более древние остатки крайне редки и включают лишь несколько фрагментов. Предположительно, во время голоцена птицы предпочитали восточное побережье Южного острова. Уорти с соавторами обнаружили остатки четырёх особей A. defossor близ Такака; намного больше остатков возрастом 13—100 тыс. лет было обнаружено в долине реки (места раскопок около северо-западного побережья Южного острова) .
Уорти и Ричард Холдавэй ( Richard N. Holdaway ) в 2002 году предположили, что птицы предпочитали селиться в кустарниках и на полях, которые покрывали восточное побережье новозеландских островов во времена голоцена. Вуд (Wood) с соавторами в 2017 году на основании того факта, что не менее 19 особей было обнаружено в долине реки Опарара на слое, расположенном непосредственно над слоем смешанного подокарпово - букового леса, посчитали, что апторнисы предпочитали сухие подокарповые леса .
По-видимому, птицы вымерли после появления в Новой Зеландии маори в XIII веке. Истребление птиц на островах совпало с потерей среды обитания и проникновением малой крысы ( Rattus exulans ) и собак. К появлению европейцев в XVII веке апторнисы уже вымерли . Боаст (Boast) полагал, что апторнисы, как и моа, вымерли около 1500 года .
Питание
Особое внимание учёных привлекал рацион апторнисов. Оуэн на основании морфологии клюва птиц построил гипотезу, что рацион включал земляных червей и личинок чешуекрылых (молей и бабочек). Он полагал, что апторнисы могли «вонзать» сильно вытянутый клюв в землю, а также копать и скрести землю лапами, на что указывает их строение. В двух почти полных скелетах из в Новой Зеландии было обнаружено 35 камней массой 16 г и 23 камня массой 8 г . Такие малые размеры камней также свидетельствуют в пользу плотоядной теории, так как у травоядных птиц камни в желудке крупнее. В пользу этой теории выступает и обнаруженный в ходе анализа изотопов больший уровень 15 N в костях апторнисов по сравнению с травоядными южным моа ( Euryapteryx curtus ), и насекомоядным Aegotheles novaezealandiae . Возможно, копаясь в гнилых брёвных и норах, апторнисы также питались более сложными беспозвоночными, включая двустворчатых , и позвоночными, например, киви . Не исключено, что в пищу попадали и морские животные, об этом свидетельствует низкий уровень 13 C .
Боаст также полагал, что апторнисы были хищниками, но не исключал, что они могли питаться падалью . Вероятно апторнисы находили добычу ночью с помощью обоняния. На это указывают большие слёзные железы .
Размножение
Среди обнаруженных остатков, относящихся к апторнисам, в различных музеях хранятся фрагменты яичной скорлупы .
Систематика
В англоязычной литературе вместо латинского Aptornis proasciarostratus принято название Adzebill (адзебилл) в знак уважения к названию, данному в работе В. Паркера: он употребил его в 1866 году по отношению к «птице с клювом в форме тесла » .
В 1848 году британский палеонтолог Гидеон Мантелл описал род Aptornis чтобы отличать остатки от больших моа ( Dinornis ) и Palapteryx (современные систематики считают этот род синонимичным Dinornis ) . Hesse в 1990 году ошибочно применил название Apterornis , которое ранее использовалось для описания других птиц, в том числе маврикийского дронта .
Боаст с соавторами предположили, что разделение на Aptornis defossor и Aptornis otidiformis произошло в плейстоцене , около 1,1 млн лет назад (0,2 — 2,3 млн лет назад) .
Учёные выделяют три вида апторнисов :
- Aptornis defossor (Owen, 1843) — Южный остров
- Aptornis otidiformis (Owen, 1849) — Северный остров
- Aptornis proasciarostratus (Worthy, 2011) — Южный остров
Сложности с определением систематического положения апторнисов связаны с их большими размерами. Ранние исследователи полагали апторнисов базальным таксоном для журавлеобразных и связывали их с вымершей нелетающей птицей из Новой Каледонии — кагу ( Rhynochetos jubatus ) , которая позднее была перенесена в отряд Eurypygiformes . Среди современных учёных такой точки зрения придерживались Брэдли Ливзи ( Bradley C. Livezey ) и Ричард Зуси ( Richard L. Zusi ), поместившие в 2007 году семейства Aptornithidae и Rhynochetidae , вместе с семейством Eurypygidae в подотряд Eurypygae .
Если предположить, что апторнис не является Eurypygiformes и не находится в близком родстве с кагу, а является родственным пастушковым (Rallidae) того же отряда журавлеобразных, то возникает вопрос, какая из современных птиц семейства наиболее близка этому виду . В 1879 году Оуэн предположил, что ближайшим родственником апторнисов является такахе ( Porphyrio hochstetteri ) — нелетающая новозеландская птица с крупным черепом, широким клювом и небольшим размером мозговой полости. В. Паркер в 1866 году предлагал отнести апторнисов к роду Notornis , к которому в то время относили такахе. Позднее птиц сравнивали с трубачами ( Psophia ) и кладой «настоящих» журавлеобразных .
В конце XX века появилась ещё одна теория, согласно которой апторнисы могут быть родственны представителям надотряда Galloanseres , включающего гусеобразных и курообразных .
Связь с кагу
В XX веке к журавлеобразным относили несколько ископаемых семейств птиц, найденных на фрагментах Гондваны , в частности в Новой Каледонии , Новой Зеландии , на Мадагаскаре и в Южной Америке . Предполагается, что их общий предок обитал на материке до его разделения. При этом апторнисов считали в близком родстве с кагу из Новой Календонии, полагая что в палеогене Новая Зеландия и Новая Каледония могли быть соединены . Ливзи с соавторами в 1998 и 2007 годы рассматривали сестринскую группу, включающую апторнисов и кагу .
Современные молекулярные исследования существенно пересмотрели систематическое положение таксонов внутри отряда журавлеобразных . В частности, кагу были признаны родственными солнечным цаплям ( Eurypyga ) и вынесены за пределы отряда . Таким образом, по мнению и морфологический анализ Ливзи и Зуси нуждается в пересмотре, так как учёные исходили из монофилии журавлеобразных . Согласно их исследованиям, опубликованным в 2019 году, апторнисы не имеют сходства ни с кагу, ни с представителями надотряда Galloanseres .
Связь с Sarothruridae
В 1997 году Houde с соавторами предположили, что апторнис является сестринским по отношению к лапчатоногим (Heliornithidae) . Позднее были обнаружены родственные связи лапчатоногих и пушистых погонышей ( Sarothrura ), в результате чего последние были выделены в отдельное семейство Sarothruridae . в 2019 году Боаст с соавторами провели более полный анализ, основанный на почти полном геноме апторнисов с Северного и Южного островов и обнаружили сестринские связи апторнисов с новым семейством . Согласно исследованиям митохондриальной ДНК, опубликованным Боастом с соавторами в 2019 году, разделение Ralloidea и Gruoidea произошло около 64 млн лет назад, появление Ralloidea — 54 млн лет назад, а отделение апторнисов от Sarothruridae — 39 млн лет назад. Такие оценки означают, что распространение Neoaves происходило в позднем меловой периоде .
Вместе с тем, морфологические исследования Муссер и Кракрафта, также опубликованные в 2019 году и включающие более сотни различных параметров, не показали однозначной связи ни в одной из групп Aptornis + M. kioloides , Aptornis +Sarothruridae и Aptornis +Sarothruridae+Heliornithidae .
Связь с трубачами
Муссер и Кракрафт в качестве основной приняли гипотезу о сестринском отношении апторнисов и трубачей, в противном случае они полагали, что апторнисы могут быть связан с «настоящими» журавлеобразными . Связь нелетающих новозеландских апторнисов со слабо летающими южноамериканскими трубачами кажется невозможной в силу большой географической удалённости регионов, которые разделяет 8—10 тысяч км. Однако подобные родственные связи были обнаружены в группе Rhynochetos + Eurypyga , где нелетающие кагу обитают в Новой Каледонии, а слабо летающие солнечные цапли — в Южной Америке .
Для определение границ, во время которых произошло разделение апторнисов и трубачей учёные рассматривали разделение Ralloidea и Gruoidea и обособление трубачей от журавлепободных птиц. По оценкам Claramunt и Кракрафта 2015 года первое случилось 56 млн лет назад, а второе — 42 млн лет назад. Prum с соавторами в том же году оценили возраст этих событий как 40 и 31 млн лет, соответственно. Схожие оценки предложил García-R с соавторами в 2014 году: отделение трубачей, по их мнению, случилось 35 млн лет назад. Claramunt и Кракрафт также предположили, что разделение кагу и солнечными цаплями произошло 33—32 млн лет назад . Учёные полагают что отделение стем-группы Rhynochetos + Eurypyga от фаэтонообразных (Phaethontiformes) случилось 61—58 млн лет назад .
Такие различия в оценках Муссер и Кракрафт объяснили различиями в используемых методах ( англ. slow nuclear, constant-rate genes ) против ( англ. mostly exonic anchored-probe dataset ) или ( англ. mitochondrial sequences ). В попытках интерпетировать эти данные Муссер и Кракрафт оценили возраст появления обоих групп Aptornis + Psophia и Rhynochetos + Eurypyga в 40—30 млн лет. Они полагают, что предки Rhynochetos + Eurypyga были широко распространены на территории Гондваны в регионах, позднее сформировавших Южную Америку, Антарктику и Австралию, в том числе Зеландию. Выводя предков «настоящих» журавлеобразных к Западной Гондване (а именно Южной Америке и соответствующей части Антарктиды), учёные полагают, что предки Aptornis + Psophia жили в том же географическом регионе, что и предки Rhynochetos + Eurypyga . Вместе с тем, учитывая возраст таксонов, не исключено, что имело место распространение не по земле, а над массивами воды; разделение биогеографических регионов произошло десятки миллионов лет назад, в то время как разделение таксонов случилось не так давно. Возможно, для разрешения этого вопроса необходимы ещё не обнаруженные более старые остатки апторнисов и журавлеобразных .
Примечания
-
Докинз Р.
Рассказ предка. Паломничество к истокам жизни. — ACT, 2021. —
ISBN 9785457889446
.
Одной из самых эффектных птиц был апторнис (Aptornis) — дальний родственник журавлей и пастушков
- ↑ , pp. 3—5.
- ↑ Boast A. P., Chapman B., Herrera M. B., Worthy T. H., Scofield R. P., Tennyson A. J. D., Houde P., Bunce M., Cooper A., Mitchell K. J. (англ.) // Diversity. — 2019. — Vol. 11. — doi : . 19 ноября 2021 года.
- ↑ Worthy T. H., Tennyson A. J. D., Scofield R. P. (англ.) // Journal of Ornithology. — 2011. — P. 669—680. — doi : .
- Parker W. K. от 17 января 2022 на Wayback Machine //The Philosophical Transactions of the Royal Society. 1866, 156. P.113–183.
- ↑ (англ.) информация на сайте Paleobiology Database . (Дата обращения: 17 декабря 2021)
- ↑ , pp. 5—7.
- ↑ Livezey B. C., Zusi R. L. (англ.) // Journal of Ornithology. — 2007. — Vol. 149. — P. 1—95. — doi : . 9 июня 2021 года.
- ↑ , pp. 28—29.
- , p. 26.
- ↑ , pp. 26—28.
Ссылки
- Musser G. M., Cracraft J. (англ.) // American museum novitates. — 2019. — P. 1—69.
|
|
|
|---|
- 2021-09-25
- 1