Целурозавры
- 1 year ago
- 0
- 0

Тираннозаври́ды ( лат. Tyrannosauridae ) — семейство тероподовых динозавров , в которое включают два подсемейства: альбертозаврины (Albertosaurinae) и тираннозаврины (Tyrannosaurinae), в свою очередь, содержащие до 13 родов, включая эпоним группы — род тираннозавр ( Tyrannosaurus ), а также трибу Alioramini . Различные источники называют разное количество таксонов, включённых в семейство; некоторые эксперты распознают только 3 рода. Все представители семейства жили на протяжении позднего мелового периода и их ископаемые остатки найдены исключительно в Азии и Северной Америке .
Хотя тираннозавриды произошли от более мелких предков, они почти всегда были крупнейшими хищниками в своих экосистемах , находясь на вершине пищевой цепи . Самым крупным видом являлся тираннозавр рекс ( Tyrannosaurus rex ) — один из крупнейших известных наземных хищников, длина которого составляла до 12,3 метра, а масса, в соответствии с современными оценками — от 8,4 до 14 тонн. Тираннозавриды были бипедальными плотоядными животными с большими черепами , полными крупных зубов. Несмотря на большие размеры, их ноги были пропорционально длинными и хорошо приспособленными для быстрого передвижения. Напротив, их передние конечности были очень маленькими и были оснащены только двумя пальцами.
Тираннозавриды известны по многочисленным почти полным скелетам, которые позволяют учёным проводить самые разнообразные исследования, касаемые биологии этих животных, такие, как онтогения , биомеханика и экология . Один из сохранившихся образцов тираннозавра рекса содержит остатки мягких тканей.

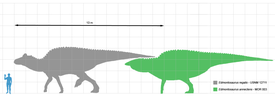
Все тираннозавриды были крупными животными. Минимальная масса для любого вида составляла 1 тонну . Длина экземпляра алиорама , найденного в 1976 году, оценивается в 5—6 метров , хоть он и считается принадлежавшим неполовозрелой особи . Альбертозавр , горгозавр и дасплетозавр достигали длины от 8 до 10 метров , в то время как тарбозавр вырастал до 12 метров . Массивный тираннозавр достигал 12,3 метра длины у самого крупного известного экземпляра (FMNH PR2081) с массой от 8,4 до 14 тонн .
Анатомия черепов представителей группы изучена хорошо, поскольку полные черепа известны у всех родов тираннозаврид. Тираннозавр, тарбозавр и дасплетозавр обладали черепами, имевшими длину более 1 метра . Взрослые особи тираннозаврид имели высокие, крепкие черепа, усиленные слитыми костями для крепости. В то же время, полости между костями и несколько окон помогали облегчить вес черепа. Многие черты черепов тираннозаврид также были обнаружены у их непосредственных предков, включая высокие предчелюстные кости и сросшиеся носовые кости .
Черепа тираннозаврид обладают множеством уникальных характеристик, включая слитые теменные кости с выраженным сагиттальным гребнем , который проходит продольно вдоль сагиттального шва и разделяет два надвисочных окна на крышке черепа. За этими окнами у представителей группы расположен высокий затылочный гребень, который также выходит из затылочных костей, но вытянут в поперечной, а не в продольной плоскости. Затылочный гребень был особенно заметно развит у тираннозавра, тарбозавра и алиорама. У альбертозавра, дасплетозавра и горгозавра были высокие гребни перед глазами, на слёзных костях , в то время как у тарбозавра и тираннозавра были чрезвычайно утолщённые заглазничные кости, формирующие за глазами гребни в форме полумесяца. Вершина морды алиорама несла ряд из шести костных гребней, сформированных носовыми костями; более низкие гребни идентифицированы у некоторых образцов дасплетозавра и тарбозавра, а также у более базального тираннозавроида аппалачиозавра .

Череп располагался на конце толстой, S-образной шеи, а длинный хвост действовал как противовес, уравновешивая тяжёлые голову и туловище, формируя центр масс в районе бёдер. Тираннозавриды известны пропорционально короткими двупалыми передними конечностями, хотя иногда встречаются остатки рудимента третьего пальца :61 .
Передвигались тираннозавриды исключительно на задних конечностях, поэтому их ноги были очень массивными. В отличие от передних, задние конечности были пропорционально длинными, длиннее, чем у почти любых прочих теропод. У подростковых и молодых особей тираннозаврид (а также некоторых базальных тираннозавроидов) длина голени была больше длины бедра, что характерно для быстро передвигающихся животных, например, орнитомимид . Пропорции ног более крупных особей характерны для медленно передвигающихся животных, но не в той степени, которая наблюдается у других крупных теропод, таких как абелизавриды или карнозавры . Третья плюсневая кость у тираннозаврид была зажата между второй и четвёртой плюсневыми костями, образую структуру, известную как арктометатарзус . Неясно, когда появилась эта черта: у ранних представителей семейства, таких, как дилун ( Dilong ), она отсутствует , но была обнаружена у более позднего аппалачиозавра . Эта структура присутствовала у троодонтид , орнитомимид и ценагнатид :61 , но её отсутствие у самых ранних тираннозаврид указывает на то, что данный признак был приобретён в результате конвергентной эволюции .
Все тираннозавриды, как и их тираннозавроидные предки, были гетеродонтными животными: их предчелюстные зубы имели в разрезе форму полукруга и были меньше по размеру, чем остальные зубы. В отличие от ранних тираннозавроидов и большинства других теропод, верхнечелюстные и нижнечелюстные зубы зрелых тираннозаврид не похожи на лезвия, а сильно утолщены и часто имеют круглое поперечное сечение, причём у некоторых видов на зубах имелась редуцированная пилообразная насечка . Количество зубов, как правило, в пределах вида было одинаковым, а у более крупных видов количество зубов меньше, чем у более мелких. Например, количество зубов у алиорама составляло от 76 до 78, в то время как тираннозавр нёс от 54 до 60 зубов в своих челюстях .
В 2001 году палеонтолог заметил, что насечка на зубах альбертозавра напоминает зубной канал, оканчивающийся полостью, называемой ампулой. Тираннозавриды использовали зубы для удержания добычи и снятия мяса с костей, поэтому, когда динозавр тянул мясо, сила натяжения могла привести к сколам на зубах животного. Наличие ампулы распределило бы эту силу по бо́льшей площади поверхности и уменьшило бы риск повреждения зуба. В одной из своих работ Эблер продемонстрировал, что стержень из плексигласа с бороздками-зарубками и просверленными отверстиями более чем на 25 % прочнее, чем с обычными надрезами. В отличие от тираннозаврид и прочих теропод, древние хищники, такие, как фитозавры и диметродоны , не имели адаптаций, препятствующих возникновению трещин в зубах .

Первые окаменелые остатки тираннозаврид были обнаружены во время экспедиций, проведённых , во время которых были найдены многочисленные разрозненные зубы. На основании этих зубов в 1856 году американский биолог Джозеф Лейди ввёл в систематику новый таксон Deinodon horridus . Первые наиболее полные остатки тираннозаврид были найдены в формации в канадской провинции Альберта . Они включали несколько черепов и почти полные скелеты. Эти остатки в 1876 году изучил Эдвард Коп , который посчитал их принадлежащими восточной разновидности тираннозавроидов — дриптозаврам . В 1905 году Генри Осборн заключил, что окаменелости из Альберты значительно отличались от дриптозаров и присвоил им новое название: Albertosaurus sarcophagus («ящер из Альберты, питающийся мясом») . В 1892 году Коп описал несколько изолированных позвонков, которым дал название Manospondylus gigas . Это открытие игнорировалось на протяжении всего XX века и вызвало дискуссии лишь в начале 2000-х годов, когда выяснилось, что материал на самом деле принадлежал тираннозавру и имеет приоритет над названием Tyrannosaurus rex .
В своей работе 1905 года, озаглавленной «Альбертозавр», Г. Осборн описал два дополнительных образца, собранных в Монтане и Вайоминге в 1902 году во время экспедиции, организованной Американским музеем естественной истории . Изначально Осборн отнёс образцы к двум разным видам: Dynamosaurus imperiosus («мощный ящер властный») и Tyrannosaurus rex («королевский ящер-тиран»). Годом позже палеонтолог заключил, что эти два образца принадлежали одному виду. Несмотря на то, что Dynamosaurus был найден первым, название Tyrannosaurus упоминалось на одну страницу раньше в статье, описывающей оба экземпляра. Поэтому, в соответствии с Международным кодексом зоологической номенклатуры , стали использовать название Tyrannosaurus .
Барнум Браун , куратор Американского музея естественной истории, продолжил собирать в Альберте образцы тираннозаврид, в том числе, он нашёл первый образец, сохранивший укороченные двупалые передние лапы, характерные для группы — в 1914 году Лоуренс Ламб назвал этот экземпляр Gorgosaurus libratus («сбалансированный свирепый ящер»). Вторая важная находка горгозавра была сделана в 1942 году в виде хорошо сохранившегося, хотя и необычно маленького, полного черепа. Только после окончания Второй Мировой войны этот образец описал Чарльз Гилмор , который назвал его Gorgosaurus lancesnis . В 1988 году этот череп повторно изучили Роберт Бэккер , Филип Карри и Майкл Уильямс и на его основе ввели в систематику новый род нанотираннус ( Nanotyrannus ) . В 1946 году советские палеонтологи обнаружили первых азиатских тираннозавридов. В 1955 году советский палеонтолог Евгений Малеев описал новые монгольские виды тираннозавра и горгозавра, а также один новый род тарбозавр ( Tarbosaurus ). Последующие исследования, однако, показали, что новый вид тираннозавра Малеева был тарбозавром на разных стадиях роста. Второй вид монгольского тираннозаврида нашёл и описал Сергей Курзанов, вид получил название Alioramus remotus («отдалённая другая ветвь»), хотя его статус истинного тираннозаврида, а не более отдалённого тираннозавроида, всё ещё остаётся спорным .
Количество описанных видов тираннозаврид и тираннозавроидов резко увеличилось в XXI веке. Это связано не только с открытием новых видов, таких как литронакс аргестес ( Lythronax argestes ) или нанукзавр Хоглунда ( Nanuqsaurus hoglundi ), но и с перенесением уже известных таксонов в группу тираннозавроидов .

В то время как остатки ранних тираннозавроидов обнаружены в трёх северных частях света , окаменелости тираннозаврид найдены исключительно в Азии и Северной Америке. Окаменелые остатки, приписываемые тираннозавридам, были обнаружены в Бразилии и даже Австралии , однако их принадлежность к данной группе оспаривается, при этом они не однозначно тираннозавровые по признакам. Австралийский экземпляр, например, представляет немногим больше, чем часть таза, и хотя он сохранил увеличенный лобковый «башмак», его принадлежность к семейству крайне спорна :98—99 . Точное время и место происхождения семейства остаются неизвестными из-за плохой сохранности среднемеловых окаменелостей на обоих континентах, хотя самые ранние известные тираннозавриды обитали в ранне кампанскую эпоху в западной части Северной Америки .
Окаменелости, принадлежавшие тираннозавридам, никогда не находили в восточной части Северной Америки, в то время как более базальные тираннозавроиды — например, дриптозавр и аппалачиозавр — сохранялись там до конца мелового периода. Это говорит о том, что тираннозавриды развивались и расселялись в западной части Северной Америки после того, как в середине мела континент раскололся пополам с образованием Западного внутреннего моря . Окаменелости тираннозаврид найдены на Аляске , через которую пролегал возможный путь расселения этих динозавров из Северной Америки в Азию . В одном кладистическом анализе обнаружена родственная связь алиорама и тарбозавра , которые образовывали уникальную азиатскую ветвь семейства . Позднее эта связь была опровергнута с открытием цяньчжоузавра и введением в систематику трибы Alioramini .

Из двух подсемейств тираннозаврины , по-видимому, были распространены шире, чем альбертозаврины . Последние неизвестны из Азии, где обитали тираннозаврины, такие, как тарбозавр и чжучэнтираннус , а также цяньчжоузавр и алиорам из трибы Alioramini. Остатки представителей обеих подсемейств присутствуют в кампанских и нижне маастрихтских отложениях Северной Америки, причём окаменелости тираннозаврин, например, дасплетозавра , присутствуют на всей западной части континента, в то время как остатки альбертозаврин — альбертозавра и горгозавра — известны только из северо-западной части континента .
К концу маастрихтской эпохи альбертозаврины, по-видимому, вымерли, а тираннозаврин тираннозавр бродил на широких пространствах от Саскачевана до Техаса . Подобная картина находит отражение и среди прочих североамериканских таксонов динозавров. Во времена кампанской — ранней маастрихтской эпох на северо-западе были распространены ламбеозавриновые гадрозавры и центрозавриновые цератопсы , в то время как хасмозаврины и гадрозаврины были распространены на юге. К концу мелового периода остатки центрозаврин уже не встречаются, а остатки ламбеозаврин редки, но гадрозаврины и хасмозаврины были распространены во всем западном регионе . В 2016 году Стив Брусатти и Томас Карр с соавторами высказали гипотезу, что в позднем маастрихте сам тираннозавр мог быть частично ответственен за вымирание других тираннозаврид на большей части запада Северной Америки. Исследование показало, что тираннозавр мог иммигрировать из Азии, а не быть автохтоном Северной Америки. Он мог являться потомком близкородственного тарбозавра, и после переселения на североамериканский континент попросту вытеснил всех прочих тираннозаврид. Данная гипотеза косвенно подтверждается тем фактом, что в пределах известных ареалов тираннозавров обычно мало или вовсе отсутствуют другие виды тираннозаврид .

Палеонтолог и его коллеги изучили рост и историю жизни тираннозаврид. С помощью гистологического анализа костей можно определить возраст, в котором умер динозавр. Можно изучить темпы роста, а результаты изобразить в виде кривых на графике. Эриксон и его коллеги выяснили, что после длительного пребывания в подростковом возрасте тираннозавры испытывали огромный всплеск роста в течение 4 лет примерно в середине жизни. Фаза быстрого роста завершалась половым созреванием , и рост уже взрослых животных значительно замедлялся. Кривая роста тираннозаврида имеет S-образную форму, с максимальной скоростью роста особей в возрасте около 14 лет .

По оценкам, самая маленькая известная особь тираннозавра ( LACM 28471, «теропод Джордана») весила всего 29,9 килограмма в возрасте 2 лет, тогда как самый большой экземпляр (FMNH PR2081, « ») в возрасте 28 лет весил 5654 килограмма; этот возраст считается близким к максимально возможному для данного вида . Подростковые особи тираннозавра весили менее 1800 килограммов до тех пор, пока им не исполнялось примерно 14 лет, после чего размер тела начинал резко увеличиваться. Во время этой фазы быстрого роста молодой тираннозавр прибавлял около 600 килограммов в год в течение следующих 4 лет. Набор массы замедлялся после 16 лет, а после 18 лет рост прекращался совсем . Например, всего 600 килограммов массы отделяли 28-летнюю «Сью» от 22-летнего канадского образца ( RTMP 81.12.1) . Это внезапное изменение в скорости роста может указывать на физическую зрелость — данная гипотеза подтверждается открытием в бедре 18-летнего тираннозавра из Монтаны (MOR 1125, также известный как «Би-рекс») . Медуллярная ткань обнаруживается только у самок во время овуляции, что указывает на то, что «Би-рекс» был репродуктивного возраста .
Другие тираннозавриды демонстрируют чрезвычайно похожие кривые роста, хотя и с более низкими темпами, соответствующими их размерам . По сравнению с альбертозавринами, дасплетозавр показывает бо́льшие темпы роста в течение периода взросления из-за более высоких показателей массы у взрослых особей. Максимальное прибавление массы у дасплетозавра составляло 180 килограммов в год при общей массе тела у взрослых особей до 1800 килограммов. Другие авторы предложили более высокую массу взрослых особей дасплетозавра; это изменило бы величину темпа роста, но не общую картину . Самый молодой известный альбертозавр, остатки которого найдены в костеносном слое Драй Айленд, погиб в возрасте 2 лет. Его длина чуть превышала 2 метра, а прижизненная масса оценивается в 50 килограммов. Окаменелости 10-метровой, 28-летней особи из того же карьера принадлежат самому крупному и пожилому из известных альбертозавров. Период стремительного роста у этих животных приходился на 12—16 лет, а ежегодная прибавка в массе составляла 122 килограмма в год до достижения 1300 килограммов у взрослых особей, что составляет примерно 1/5 массы тираннозавра. Горгозавры в период роста набирали примерно 110 килограммов в год; столько же набирали альбертозавры .

Конец фазы быстрого роста предполагает начало половой зрелости у альбертозавра, хотя в течение жизни животных рост продолжался более медленными темпами . Процесс полового созревания во время фазы активного роста, очевидно, характерны как для мелких , так и крупных динозавров, а также для крупных млекопитающих, таких как люди и слоны . Эта картина относительно раннего полового созревания разительно отличается от картины, наблюдаемой у птиц, половая зрелость которых наступает после прекращения роста .
Проанализировав все образцы в каждой возрастной группе, Эриксон и его коллеги смогли сделать выводы об истории жизни в популяциях тираннозаврид. Их анализ показал, что в то время как окаменелые остатки неполовозрелых особей встречаются редко, остатки подростков в активной фазе роста и взрослых животных распространены гораздо чаще. Более половины известных экземпляров тираннозавров умерли в течение 6 лет после достижения половой зрелости. Такая картина наблюдается у других тираннозаврид, а также у современных крупных, долго живущих птиц и млекопитающих. Эти виды характеризуются высокой детской смертностью, за которой следует относительно низкая смертность среди неполовозрелых особей. Уровень смертности вновь увеличивался среди половозрелых животных, частично из-за ран, полученных при размножении. Несмотря на возможные неточности при извлечении или обработке образцов, Эриксон высказал предположение, что такая разница могла быть обусловлена низкой смертностью среди ювенильных особей определённого размера, что также наблюдается у некоторых современных крупных млекопитающих, например, слонов. Эта низкая смертность, возможно, была вызвана отсутствием естественных врагов, поскольку к двум годам тираннозавры превосходили размером всех прочих хищников. Палеонтологи не обладают необходимым количеством окаменелых остатков дасплетозавра для подобного анализа, но Эриксон полагет, что похожая тенденция применима и к этим животным .
Тираннозавриды проводили половину своей жизни неполовозрелыми, прежде чем начинали стремительно расти, достигая максимального размера за несколько лет . Это, наряду с полным отсутствием промежуточных по размеру хищников между огромными взрослыми тираннозавридами и другими мелкими тероподами, позволяет предположить, что экологические ниши мелких хищников могли быть заполнены ювенильными тираннозавридами. Такая картина наблюдается у современных комодских варанов , чьи детёныши начинают как насекомоядные, обитающие на деревьях, но постепенно вырастают в крупных хищников, способных убивать других позвоночных . Например, окаменелые образцы альбертозавров были обнаружены в виде скоплений костей, принадлежавших особям разного возраста .
Способности тираннозаврид к передвижению лучше всего изучать на образцах тираннозавра. Здесь есть два важных вопроса: как быстро он мог поворачиваться и какова была его максимальная прямолинейная скорость. Тираннозавр был медлительным при повороте: считается, что он поворачивался на 45° за одну или две секунды — на это человеку, вертикально ориентированному и лишённому хвоста, потребовались бы доли секунды . Причиной такой медлительности являлась инерция вращения животного, поскольку бо́льшая часть его массы находилась на некотором расстоянии от центра тяжести , как у человека, несущего бревно .

Оценки максимальной скорости тираннозавра разнятся: в среднем скорость оценивается в 11 метров в секунду, некоторые называют цифру 5—11 м/с (18—39,6 км/ч), и лишь немногие — до 20 м/с (72 км/ч). Исследователи вынуждены полагаться на различные методы оценки, потому что, хотя существует много следов, оставленных крупными тероподами, до сих пор не найдено ни одного следа бегущего крупного теропода, а это может указывать на то, что они не бегали .
В 1993 году Джек Хорнер и Дон Лессем предположили, что тираннозавр был медлительным животным и, вероятно, не мог бегать . Тем не менее, в 1996 году Томас Хольц утверждал, что тираннозавриды и их родственники были самыми быстрыми крупными тероподами . П. Кристиансен в 1998 году подсчитал, что кости ног тираннозавра не были сильнее костей ног слонов, чья скорость ограничена и не предполагает быстрого бега. Он предположил, что максимальная скорость, с которой мог двигаться тираннозавр, составляла 11 м/с (39,6 км/ч), то есть со скоростью спринтера . Джеймс Фарлоу и его коллеги в 1995 году утверждали, что тираннозавр получил бы критическую или даже смертельную травму, если бы он упал при быстром движении, поскольку его туловище врезалось бы в землю с ускорением в 6 g (то есть в 6 раз больше ускорения силы тяжести, около 60 м/с), и его крошечные ручки не смогли бы смягчить удар . Тем не менее, известно, что жирафы скачут со скоростью до 50 км/ч, несмотря на риск сломать ногу или хуже, что может быть смертельным даже в безопасной среде, такой как зоопарк . Таким образом, вполне возможно, что тираннозавр также двигался быстро, когда это было необходимо; это утверждение справедливо и для аллозавра . Самые последние исследования локомоции тираннозавра выявляют диапазон скорости 17—40 км/ч — от ходьбы до медленного бега . При компьютерном моделировании, проведённом в 2007 году на основе данных, полученных непосредственно из окаменелостей, была выявлена максимальная скорость движения тираннозавра, равная 8 м/с (28,8 км/ч) . Правда, окаменелости, возможно, принадлежали ювенильной особи .
Судя по окаменевшим отпечаткам ног в районе Гранд-Прери в Канаде возрастом 72,5 млн л. н., тираннозавриды с увеличением размеров и массы тела претерпевали снижение относительной манёвренности и скорости передвижения, а значит юные тираннозавриды были быстрее и подвижнее своих родителей .


Продолжительные дебаты в палеонтологическом сообществе посвящены степени и характеру кожного покрова тираннозаврид. Длинные нитевидные структуры сохранились вместе со скелетными остатками многочисленных целурозавров раннемеловой формации и других близлежащих геологических формаций из Ляонина ( Китай ) . Эти нити обычно интерпретируются как «протоперья», гомологичные разветвлённым перьям, обнаруженным у птиц и некоторых нептичьих теропод , хотя существуют и другие гипотезы . В 2004 году был описан новый таксон — дилун ( Dilong ), который имеет первые доказательства наличия протоперьев у тираннозавроидов. Подобно пуховым перьям современных птиц, протоперья дилуна были разветвлёнными, но не контурными и, возможно, использовались для теплоизоляции . Открытие и описание в 2012 году 9-метрового тираннозавроида ютираннуса ( Yutyrannus ) может означать, что и крупные тираннозавриды были покрыты перьями .
Исходя из принципа , было предсказано, что тираннозавриды также могут обладать перьевым покровом. Тем не менее, исследование, опубликованное группой учёных в журнале « Biology Letters » в 2017 году, описало отпечатки кожи тираннозаврид, собранные в Альберте , Монтане и Монголии , которые принадлежали 5 родам динозавров: тираннозавру, альбертозавру, горгозавру, дасплетозавру и тарбозавру . Хотя отпечатки кожи невелики, они представляют части почти всего тела животного, кроме головы: брюха, грудной области, таза, хвоста и шеи. Образцы содержат отпечатки узкой, тонкой, неперекрывающейся чешуи (которую соавтор статьи Скотт Персонс сравнивает с чешуёй на боках крокодила ) и не несут никаких следов оперения. Базовая текстура состоит из крошечных «основных чешуек» диаметром 1—2 миллиметра, а некоторые оттиски показывают следы 7-миллиметровых «отличительных чешуек», расположенных среди них. Кроме того, чешуйки можно разглядеть в отпечатках лап тираннозаврид , а черепа тираннозаврид содержат остеологические корреляты чешуи .
Группа палеонтологов под руководством Филиппа Белла выполнила реконструкцию предкового таксона тираннозаврид, основываясь на известных данных о покрове этих животных. Несмотря на 89 % вероятность наличия перьев у предка тираннозаврид, учёные определили, что вероятность появления чешуйчатых тираннозаврид составляет 97 %. Данные «представляют убедительные доказательства полного чешуйчатого покрова у тираннозавра», — написал Белл и его команда. Палеонтологи также признали, что оперение могло присутствовать в грудной области животного, отпечатки которой не были обнаружены .
Эту статью подвергли сомнению некоторые палеонтологи, такие как Андреа Кау и Томас Хольц, которые указали, что перья у тероподовых динозавров могут расти на тех же частях тела, что и чешуя (как у юравенатора ), и, таким образом, наличие чешуи фактически не указывает на отсутствие перьев на этой части тела . Кроме того, перья являются тонкими, непрочными структурами, которые могли легко отделяться от тела при разложении. Палеонтолог и палеоиллюстратор заявил, что этих динозавров следует изображать более чешуйчатыми, чем оперёнными, — в том виде, к которому люди привыкли. Признавая роль, которую играет тафономия в интерпретации кожи динозавра, Уиттон отметил, что оттиски чешуек имеют высокое качество, одинаковы по форме на всех участках и не показывают явных свидетельств отпечатков волокон или пространства для крепления нитевидных волокон, несмотря на сохранившиеся отпечатки чешуек размером меньше 1 миллиметра. Если бы при разложении тела образец был значительно искажён, можно было ожидать большей деформации кожи образцов .
Причины потери тираннозавридами перьевого покрова пока не ясны. Постепенную потерю перьев можно наблюдать и у других групп динозавров, таких как птицетазовые , которые утеряли нитевидные волокна и вновь обрели чешуйки. Одной из возможных причин называют гигантизм животных, однако Фил Белл отметил, что оперённый ютираннус размером сравним с горгозавром и альбертозавром. По мнению учёного, проблема в том, что в одно и то же время существовали тираннозавриды, покрытые перьями, и тираннозавриды, покрытые чешуёй, а причина такой разницы в покровах тела неизвестна .

Глазницы тираннозавра были развёрнуты вперёд, давая ему бинокулярное зрение , которое было немногим лучше, чем у современных ястребов . В то время как у хищных теропод в целом бинокулярное зрение было непосредственно перед глазами, у тираннозаврид была значительно бо́льшая область наложения взгляда. Джек Хорнер также отметил, что эволюция тираннозаврид неуклонно улучшала бинокулярное зрение. Трудно понять, как естественный отбор способствовал бы этой долгосрочной тенденции, если бы тираннозавриды были исключительно падальщиками — им, как известно, не требуется восприятие глубины, которое обеспечивает стереоскопическое зрение . У современных животных бинокулярное зрение встречается в основном у хищников (главное исключение составляют приматы , которым оно необходимо для прыжка с ветки на ветку). В отличие от тираннозавра, у тарбозавра был более узкий череп, типичный для других тираннозаврид, у которых глаза были обращены преимущественно вбок. Всё это говорит о том, что тарбозавр полагался больше на обоняние и слух, чем на зрение . У горгозавра глазницы были круглыми, а не овальными или в виде замочной скважины, как у других тираннозаврид . У дасплетозавра это был высокий овал, по форме средний между круглой глазницей горгозавра и «замочной скважиной» тираннозавра .
Подробное исследование, проведённое Томасом Карром и его коллегами в 2017 году, посвящённое сравнению костяных структур дасплетозавра и крокодила, выявило, что у тираннозаврид были большие, плоские чешуйки на морде. В центре этих чешуек располагались небольшие кератинизированные участки. У крокодилов такие участки покрывают пучки сенсорных нейронов, при помощи которых эти рептилии могут обнаруживать механические, термические и химические раздражители . Учёные предположили, что тираннозавриды, вероятно, также имели пучки сенсорных нейронов под чешуйками в передней части морды и, возможно, использовали их для распознавания объектов, измерения температуры гнёзд и осторожного обращения с и детёнышами .

Костные гребни встречаются на черепах многих теропод, в том числе, тираннозаврид. Носовые кости алиорама несли ряд из 5 костяных выпуклостей. Похожий ряд выпуклостей присутствует на черепе аппалачиозавра , а также у некоторых экземпляров альбертозавра, дасплетозавра и тарбозавра . У альбертозавра, горгозавра и дасплетозавра перед каждым глазом на слёзной кости виден выдающийся рог. Слёзный рог отсутствует у тарбозавра и тираннозавра, у которых вместо этого есть гребень в форме полумесяца за каждым глазом на заглазничной кости. Все эти гребни могли использоваться тираннозавридами для демонстрации, распознавания своего вида или при брачном ухаживании .
Долгое время считалось, что тираннозавр, как и большинство динозавров, обладал эктотермическим (« холоднокровным ») метаболизмом рептилий. Однако, некоторые учёные, такие, как Джон Остром и Роберт Бэккер бросили вызов этой гипотезе в первые годы « », начавшегося в конце 1960-х годов . Сам тираннозавр рекс был эндотермическим (теплокровным) животным, что подразумевало очень активный образ жизни :Глава 10 . С тех пор несколько палеонтологов пытались определить способность тираннозавра регулировать температуру своего тела. Гистологическое подтверждение высоких темпов роста у молодых особей, сопоставимых с таковыми у млекопитающих и птиц, может говорить в пользу гипотезы о быстром метаболизме. Кривые роста показывают, что, как и у млекопитающих и птиц, рост тираннозавра ограничивался в основном неполовозрелыми животными; рост не был непрерывным, как у большинства других позвоночных . Была указана разница температур между позвонками туловища и голенью: 4—5 °C. О таком небольшом интервале между телом и конечностями заявили палеонтолог Риз Баррик и геохимик Уильям Шауэрс, чтобы показать, что тираннозавр поддерживал постоянную температуру тела ( гомойотермию ) и что он обладал метаболизмом где-то между экзотермическими рептилиями и эндотермическими млекопитающими . Позже они обнаружили аналогичные результаты в образах гиганотозавров , которые жили на другом континенте и на десятки миллионов лет раньше . Однако, даже если тираннозавр действительно демонстрирует гомойотермию, это не обязательно означает, что он был эндотермическим животным. Такая терморегуляция также может быть объяснена гигантеротермией, как у некоторых современных морских черепах .

В формации горгозавр жил рядом с более редким тираннозаврином дасплетозавром. Это один из немногих примеров сосуществования двух родов тираннозаврид. В современных экосистемах хищники одинакового размера делят разные экологические ниши анатомическими, поведенческими или географическими различиями, которые ограничивают конкуренцию. Неясно, какие именно ниши делили тираннозавриды Дайносор Парка . В 1970 году американский палеонтолог Дейл Рассел выдвинул гипотезу, что более распространённый горгозавр активно охотился на быстроногих гадрозаврид , в то время как более редкие и сильные цератопсы и анкилозавры являлись добычей более крепкого десплетозавра . Тем не менее, образец дасплетозавра OTM 200 из формации сохранил переваренные окаменелые остатки подростковой особи гадрозаврида в области кишечника . Фоссилии горгозавров чаще всего встречаются в северных формациях, таких как Дайносор Парк, а виды дасплетозавров более распространены на юге. Такая же картина наблюдается у других групп динозавров. Хасмозаврины и гадрозаврины также были более распространены в формации Ту-Медисин в Монтане и на юго-западе Северной Америки на протяжении кампанской эпохи , в то время как центрозаврины и ламбеозаврины преобладали в северных широтах. Томас Хольц предположил, что эта модель указывает на общие экологические предпочтения между тираннозавринами, хасмозавринами и гадрозавринами. В конце поздней маастрихтской эпохи тираннозаврины, такие как тираннозавр, гадрозаврины, такие как эдмонтозавр , и хасмозаврины, такие как трицератопс , были широко распространены в западной части Северной Америки, в то время как альбертозаврины и центрозаврины вымерли, а ламбеозаврины были редки .
Имеются некоторые свидетельства социального поведения тираннозаврид. Например, экземпляр тираннозавра, известный как « », по-видимому, умер от сильного укуса в голову, который мог нанести только другой тираннозавр . Исследователи сообщали о скелетах детёныша и подростка тираннозавров, найденных в том же карьере, что и «Сью». Эти находки были использованы для подтверждения гипотезы о существовании у тираннозавров социальных групп . В то время как нет никаких доказательств стайного поведения у горгозавров, свидетельства подобного поведения у альбертозавров и дасплетозавров имеются .
Образец молодого дасплетозавра из формации Дайносор Парк (TMP 94.143.1) несёт следы укусов на морде, которые были нанесены другим тираннозавридом. Следы укуса зажили, показывая, что животное пережило травму. У взрослого дасплетозавра из той же локации (TMP 85.62.1) также обнаруживаются следы укуса тираннозаврида, что свидетельствует о том, что атаки на лицо не ограничивались молодыми животными. Хотя укусы, возможно были нанесены другими видами, всё же внутривидовая агрессия распространена среди хищников. Следы укусов на морде также наблюдаются у других представителей семейства, таких как горгозавр и тираннозавр, а также среди представителей других групп, таких как синраптор ( Sinraptor ) и Saurornitholestes . Даррен Танке и Фил Карри предполагают, что укусы связаны с внутривидовой конкуренцией за территорию или ресурсы или за доминирование в социальной группе .

Доказательства того, что дасплетозавры жили социальными группами, получены из костеносного горизонта формации Ту-Медисин в Монтане. Костеносный слой содержит окаменелые остатки трёх дасплетозавров: крупного взрослого, мелкого подростка и особь среднего размера. В этой же локации сохранились остатки по крайней мере пяти гадрозавров . Геологические данные указывают на то, что фоссилии не были принесены сюда водными потоками, а что все животные были захоронены в этом месте одновременно. Остатки гадрозавров разрозненны и несут множество следов от зубов дасплетозавра — это указывает на то, что хищник питался мясом гадрозавров перед своей гибелью. Причина смерти неизвестна. Ф. Карри предположил, что дасплетозавры формировали стаи, однако этого нельзя утверждать с полной уверенностью . Другие учёные относятся скептически к предположениям о формировании социальных групп у дасплетозавров и прочих крупных теропод : Брайан Роуч и Дэниэл Бринкман предположили, что социальное взаимодействие дасплетозавров напоминало таковое у современных комодских варанов , где особи не объединяются друг с другом, поедая трупы, а чаще нападают друг на друга и даже проявляют каннибализм .
Костеносный слой Драй-Айлэнд, обнаруженный Барнумом Брауном и его командой, содержал окаменелые остатки 22 альбертозавров — самое большое количество меловых теропод, найденных в одной локации, и второе по величине среди всех теропод с учётом скопления остатков юрских аллозавров в карьере Кливленд-Ллойд в штате Юта . Захороненная здесь группа состояла из одной очень старой особи; 8 взрослых животных возрастом от 17 до 23 лет; 7 взрослых животных, проходивших фазу быстрого роста в возрасте от 12 до 16 лет; и 6 непловозрелых динозавров возрастом от 2 до 11 лет, которые не достигли фазы быстрого роста . Полное отсутствие поблизости остатков растительноядных динозавров и одинаковая степень сохранности фоссилий позволили палеонтологу Филу Карри заключить, что местность не являлась ловушкой для хищников, как смоляные ямы Ла Брея в Калифорнии, и что все сохранившиеся животные погибли в одно время. Карри утверждал, что такое захоронение указывает на стайный образ жизни альбертозавров . Другие учёные скептически относятся к данной гипотезе, считая, что животных могли собрать вместе засуха, наводнение и другие причины .
Однако, существуют доказательства, хоть и противоречивые, подтверждающие гипотезу социального поведения некоторых тираннозаврид. В расположенной в Британской Колумбии формации есть цепочка следов, оставленных тремя разными тираннозавридами (названных ихновидом Bellatoripes fredlundi ). Исследование тропы не выявило никаких признаков того, что один динозавр оставил следы после того, как прошёл другой; цепочки следов выглядят так, как если бы животные передвигались группой. Дальнейшие исследования показали, что тираннозавриды двигались со скоростью от 6,4 до 8,2 км/ч и, вероятно, имели высоту бедра от 2,13 до 2,74 метра. Поскольку из этой формации известно три разных рода тираннозаврид (альбертозавр, дасплетозавр и горгозавр), неизвестно, какие именно тираннозавриды оставили следы .
Следы зубов тираннозаврид являются наиболее часто сохраняющимися следами питания плотоядных динозавров. Отметины от их зубов находили на костях цератопсов , гадрозавров и других тираннозаврид. Кости тираннозаврид со следами зубов составляют примерно 2 % всех известных ископаемых с отметинами зубов . Тираннозавриды использовали зубы для отрывания мяса от туши, а не отрезали, как ножом. Характер износа зубов указывает на то, что представители этого семейства при отрывании мяса сильно трясли головой .

Некоторые исследователи считают, что альбертозавры охотились стаями, и предположили, что более молодые члены стаи загоняли жертву к более взрослым, которые были крупнее и мощнее, но также и медленнее . Помимо этого, молодые особи могли вести образ жизни, отличный от образа жизни взрослых, заполняя экологические ниши между крупными тероподами и мелкими, самые большие из которых были на два порядка легче взрослого альбертозавра . Эти гипотезы нельзя проверить, поскольку неопровержимые доказательства, указывающие на поведение динозавров, сохраняются в летописи окаменелостей чрезвычайно редко. Фил Карри лишь предполагал, что дасплетозавры собирались в стаи для охоты, однако и этого нельзя утверждать с уверенностью. Нет никаких доказательств стайного поведения у горгозавра .
Спор о том, был тираннозавр активным охотником или только падальщиком, столь же стар, как и спор о его локомоции. Канадский палеонтолог Лоуренс Ламб , в 1917 году описавший хорошо сохранившийся скелет горгозавра, заключил, что горгозавр, а следовательно, и его близкий родственник тираннозавр, были чистыми падальщиками, поскольку зубы горгозавра были почти не изношенными . Впоследствии этот аргумент не воспринимался всерьёз, поскольку, как выяснилось, зубы теропод регулярно менялись. Со времени первого открытия тираннозавра большинство учёных сошлись во мнении, что он был хищником, хотя, подобно современным крупным хищникам, он мог питаться и падалью или отобрать пищу у другого хищника, если бы у него была такая возможность .
Известный эксперт по гадрозаврам Джек Хорнер в настоящее время является основным сторонником гипотезы, что тираннозавр был исключительно падальщиком и вообще не занимался активной охотой . Хорнер представил несколько аргументов в пользу своей гипотезы. Присутствие больших обонятельных луковиц и обонятельных нервов предполагает наличие высокоразвитого обоняния для поиска мёртвых тел по запаху. Зубы могли раздавить кость и, следовательно, ящер мог извлечь как можно больше пищи ( костного мозга ) из остатков туши — обычно наименее питательных частей. По крайней мере, некоторые из его потенциальных жертв могли двигаться быстро, в то время как есть свидетельства того, что тираннозавр ходил, а не бегал .
Другие данные свидетельствуют об охотничьем поведении тираннозавра. Глазницы животного были развёрнуты вперёд, давая ему возможность стереоскопического зрения. Нанесённый тираннозаврами урон был обнаружен на скелетах гадрозавров и трицератопсов , которые, по-видимому, пережили первоначальные атаки . Некоторые исследователи утверждают, что, если тираннозавр был падальщиком, то в позднемеловых экосистемах Америки и Азии должен был существовать другой сверххищник, главной добычей которого были маргиноцефалы и орнитоподы . Другие тираннозавриды разделяют столько общих характеристик с тираннозавром, что ни один из них не подходит на эту роль и только мелкие дромеозавриды остаются в качестве возможных главных хищников. В свете этого приверженцы гипотезы тираннозавров-падальщиков предположили, что размер и сила этих животных позволяла им отбирать добычу у мелких хищных видов .
В 2023 году был описан образец молодого горгозавра с содержимым желудка in situ , содержащим двух неповрежденных молодых особей возрастом около года. На момент смерти горгозавр был неполовозрелым, возрастом 5—7 лет, его длина составляла около 4 метров, а масса — около 335 килограммов. Он намного крупнее двух молодых особей Citipes , которые весили 9—12 килограммов, что противоречит предположению, что тираннозавриды питались добычей своего размера, когда они достигли массы 16—32 кг, что указывает на то, что молодые тираннозавриды всё ещё потребляли гораздо меньшую добычу после превышения определённого порога размера. Обнаружение этого экземпляра показало, что тираннозавриды, вероятно, не охотились стаями, состоящими из животных нескольких поколений, поскольку размер их добычи слишком мал, чтобы их можно было разделить с сородичами. Это также прямое диетическое свидетельство, которое подкрепляет теорию «онтогенетического сдвига в рационе» тираннозаврид, как ранее предполагалось с помощью экологического моделирования и анатомических особенностей среди разных возрастных групп. В полости желудка тираннозаврид присутствовали только остатки задних конечностей и хвостовых позвонков молодых Citipes , что позволяет предположить, что молодой горгозавр, возможно, потреблял преимущественно мускулистые задние конечности .
Изначально для этого семейства Эдвард Коуп в 1866 году ввёл в систематику название Deinodontidae , которое использовалось вплоть до переименования его в Tyrannosauridae в 1960-х годах . Типовым родом для семейства Deinodontidae являлся , названный и описанный по изолированным зубам из Монтаны . Однако, в обзоре 1970 года, посвящённом североамериканским тираннозавроидам, пришёл к выводу, что род Deinodon не является валидным , и использовал название Tyrannosauridae вместо Deinodontidae, заявив, что это соответствует правилам МКЗН .
Эпоним семейства — род тираннозавр ( Tyrannosaurus ) назвал и описал Генри Осборн в 1905 году вместе с семейством тираннозаврид (Tyrannosauridae). Название происходит от греч. τυραννος/tyrannos — тиран, и σαυρος/sauros — ящер. На распространённый суффикс лат. -idae заканчиваются названия зоологических семейств , который произошёл от греч. -ιδαι/-idai , означающего множественное число .

Согласно линнеевской систематике , тираннозавриды — это семейство , находящееся в составе надсемейства тираннозавроидов отряда ящеротазовых .
Тираннозаврид разделяют на 2 подсемейства : альбертозаврины (Albertosaurinae), куда включают североамериканские роды альбертозавр и горгозавр , и тираннозаврины (Tyrannosaurinae), в состав которых входят дасплетозавр , тератофоней , бистахиэверсор , тарбозавр , нанукзавр , чжучэнтираннус и сам тираннозавр . Некоторые авторы относят вид Gorgosaurus libratus к роду альбертозавр (под названием Albertosaurus libratus ), а вид Tarbosaurus bataar — к роду тираннозавр , в то время как другие исследователи предпочитают выделять их в отдельные роды . Альбертозаврины характеризуются более лёгким телосложением, более низкими черепами и пропорционально более длинными голенями, чем у тираннозаврин . У тираннозаврин сагиттальный гребень теменных костей протягивается до лобной кости . В своей работе 2014 года группа китайских палеонтологов под руководством включила роды альбертозавр и горгозавр непосредственно в семейство тираннозаврид, а не в подсемейство альбертозаврин , однако такой систематики не придерживаются авторы более поздних работ :69 . В той же статье Люй и его коллеги описали новую группу тираннозаврид — трибу Alioramini , содержащую роды цяньчжоузавр и алиорам . Филогенетический анализ группы показал, что она находилась у основания тираннозаврин . Некоторые авторы, такие как Джордж Ольшевский и Трейси Форд, создали другие группы или трибы для различных комбинаций тираннозаврид . Однако они не были определены филогенетически и обычно включали роды, которые в настоящее время считаются синонимами других родов или видов .
Помимо этого, в систематику вводились семейства на основе более фрагментарных остатков, в том числе Aublysodontinae и Deinodontinae. Однако, роды и Deinodon ныне считаются nomina dubia , а потому их и названные на их основе подсемейства обычно исключают из таксономии тираннозаврид. Ещё один тираннозаврид, Raptorex , изначально описанный как более примитивный тираннозавроид, вероятно, представляет собой подростковую особь тираннозаврина, похожего на тарбозавра. И, поскольку он известен только по образцу подростковой особи, его название также считается сомнительным .
С появлением в палеонтологии позвоночных филогенетической таксономии семейству тираннозаврид было дано несколько чётких определений. Впервые это сделал Пол Серено в 1998 году, который определил тираннозаврид как всех тираннозавроидов, которые ближе к тираннозавру, чем к алектрозавру , Aublysodon или нанотираннусу . Тем не менее, нанотираннуса иногда считают неполовозрелым тираннозавром, а род Aublysodon считается сомнительным и непригодным для использования в определении клады .
В 2001 году американский палеонтолог опубликовал кладистический анализ тираннозаврид. Он заключил, что существует два подсемейства: более примитивные Aublysodontinae, характеризующиеся предчелюстными зубами без зазубрин, и тираннозаврины. К семейству Aublysodontinae Хольц отнёс Aublysodon , « Aublysodon из формации Киртланд» и алектрозавра. Палеонтолог также обнаружил некоторые синапоморфии тираннозаврид у сиамотирана ( Siamotyrannus ), которые, однако, «лежали вне семейства» .
Позже в той же статье Хольц предложил определять тираннозаврид как «всех потомков самого последнего общего предка тираннозавра и Aublysodon ». Также учёный подверг критике определения, ранее предложенные другими исследователями, например, одно из предложенных Полом Серено: «все тираннозавроиды, которые ближе к тираннозавру, чем к алектрозавру, Aublysodon или нанотираннусу». Хольц заметил, что поскольку нанотираннус, вероятно, был ошибочно идентифицированным молодым тираннозавром, предложенное Серено определение будет содержать само семейство тираннозаврид в качестве подтаксона рода тираннозавр. Помимо этого, его предложенное определение подсемейства тираннозаврин будет ограничено одним тираннозавром .
В 2003 году Кристофер Броху включил в семейство тираннозаврид альбертозавра, алектрозавра, алиорама, дасплетозавра, горгозавра, тарбозавра и тираннозавра . Хольц сделал повторное определение группы в 2004 году, использовав почти все вышеперечисленные таксоны в качестве спецификаторов, за исключением алиорама и алектрозавра, которых его анализ не смог уверенно определить. Однако в той же статье учёный также дал совершенно другое определение, включая всех теропод, более тесно связанных с тираннозавром, чем с эотираннусом . Самым последним определением было определение Серено в 2005 году, в котором тираннозавриды определялись как наименее инклюзивная клада, содержащая альбертозавра, горгозавра и тираннозавра .
Кладистический анализ филогении тираннозаврид часто обнаруживает, что тираннозавр и тарбозавр являются близкородственными таксонами, а дасплетозавр — более базальным , чем любой из них. Тесную связь между тираннозавром и тарбозавром поддерживают многочисленные особенности черепной морфологии, включая рисунок швов между определёнными костями, наличие серповидного гребня на заглазничной кости и очень глубокая верхняя челюсть с заметной кривизной нижнего края . Альтернативная гипотеза была представлена в исследовании Фила Карри и его коллег в 2003 году, в котором была обнаружена слабая обоснованность дасплетозавра в качестве базального члена клады, содержащей также азиатских алиорама и тарбозавра, на основании отсутствия костного зубца, соединяющего носовые и слёзные кости. В этом же исследовании алиорам был найден в качестве ближайшего родственника тарбозавра на основании сходной схемы распределения напряжения в черепе .
В связанном исследовании был отмечен механизм смыкания нижней челюсти , общий для двух родов . В отдельной статье Карри отметил возможность того, что алиорам может представлять подростковую особь тарбозавра, но заявил, что гораздо более высокое количество зубов и более заметные носовые гребни у алиорама предполагают, что это был отдельный род. Помимо этого, Карри обращает внимание на большое количество зубов у нанотираннуса и так же относит его к отдельному роду динозавров , не считая его неполовозрелым тираннозавром, как полагают большинство экспертов . Тем не менее, обнаружение и описание цяньчжоузавра показывает, что алиорам не являлся близким родственником тарбозавра, а принадлежал к недавно описанной трибе тираннозавридов — Alioramini . С открытием цяньчжоузавра стало ясно, что подобные длинноносые тираннозавриды были широко распространены по всей Азии и могли жить бок о бок, разделяя различные экологические ниши с более крупными и крепкими тираннозавринами, охотясь на другую добычу .
Кладограмма на основе филогенетического анализа, выполненного командой палеонтологов под руководством Стивена Брюссатта и Томаса Карра в 2016 году .
| Tyrannosauridae |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Кладограмма филогенетических связей внутри семейства тираннозаврид 2020 года .
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||
|
Базальные
( базальные тираннозавры ) |

|
||||||||
| Proceratosauridae | |||||||||
| ? |
|
||||||||
| Tyrannosauridae |
|
||||||||