

Орхидные
- 1 year ago
- 0
- 0

Орхи́дные , или Ятры́шниковые , также Орхиде́и ( лат. Orchidáceae ) — крупнейшее семейство однодольных растений. Более чем для 10 % представителей семейства характерен CAM-фотосинтез .
Орхидные — древнее семейство, появившееся в позднемеловую эпоху .
Название «орхидея» произошло от др.-греч. ὄρχις — яичко (человека или другого млекопитающего ) из-за формы корневища.
Классификация орхидных основана главным образом на строении их колонки и характера расположения пыльника и рыльца . Новую систему орхидных разработал американский учёный Р. Л. Дресслер : он разделяет семейство Орхидные на 5 подсемейств , 22 трибы и 70 подтриб .
|
|||||||||||||||||||||||||
В наше время орхидные найдены на всех континентах , кроме Антарктиды . Большинство видов сосредоточено в тропических широтах . Здесь, в областях с коротким сухим сезоном и высоким уровнем осадков они находят наиболее благоприятные условия для своего роста. Своеобразие флоры орхидных на разных континентах — характерная черта их распространения.
Орхидные умеренных широт — многолетние наземные травы с подземными корневищами или клубнями , а в тропиках шире всего представлены эпифитные орхидные .
В умеренном поясе флора орхидных гораздо беднее, чем в тропических широтах. На умеренные широты Северного полушария приходится только 75 родов (10 % от общего количества) и 900 видов (4,5 %). Ещё меньше — 40 родов и 500 видов — встречаются в южном умеренном поясе.
Для территории России, вместе с республиками бывшего Советского Союза, приводится 419 видов, относящихся к 49 родам .
Листья орхидных простые и очерёдные.
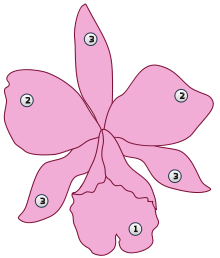
Цветки , как правило, собраны в кистевидные или колосовидные соцветия , реже цветки одиночные ( венерин башмачок , каттлея ). Цветки трёхчленные, с двойным околоцветником . Чашелистики одинаковые, а из трёх лепестков средний (морфологически верхний) сильно отличается от остальных, образуя губу . Губа нередко имеет длинный вырост назад, заполненный нектаром, — шпорец . Поскольку для опыления наиболее удобно положение «губой вниз», у наземных орхидных в процессе развития происходит перекручивание ( ресупинация ) цветка. Эпифитным орхидным, имеющим свешивающиеся соцветия, ресупинация не требуется .
Почти от всех известных цветковых растений представители семейства отличаются срастанием нитей
тычинок
, которых может быть одна, две или очень редко три, со столбиком
гинецея
в так называемую
колонку
, или гиностемий, — такое специфическое образование встречается, помимо орхидных, только у представителей семейства
Раффлезиевые
(
Rafflesiaceae
).
Формула цветка
:
![]()
Для семейства характерны значительная
редукция
андроцея
; слияние пыльцевых зёрен в гнёздах
пыльника
в
поллинии
, которые достигают очень сложного строения; резкая
зигоморфия цветка
обычно при сохранении трёх листочков наружного и трёх листочков внутреннего кругов
околоцветника
или при их частичном или полном срастании; одногнёздная или очень редко трёхгнёздная
завязь
, состоящая из трёх
плодолистиков
. Завязь нижняя, содержит очень большое число мелких семяпочек.

Механизмы опыления у орхидных отличаются большим разнообразием и оригинальностью. Как правило, целью опыления является прикрепление к насекомому-опылителю поллиниев, которые затем переносятся на другие цветки. У башмачков губа устроена по типу ловушки, попав в которую насекомое невольно производит опыление. У ятрышников и любок насекомое в поисках нектара натыкается на липкие ножки поллиниев, которые прикрепляются к его голове в виде рожек. Эти рожки за несколько минут загибаются так, что при следующем посещении цветка попадают точно на рыльце пестика . У офрис цветки имитируют вид и запах самок одиночных пчёл, привлекая в качестве опылителей самцов этих насекомых. У тропических поллинии выстреливаются в сторону насекомого-опылителя, как только оно задевает за выросты колонки. Цветки некоторых орхидей дрожат при порыве ветра, имитируя насекомых-жертв для опыляющих их цветки насекомых-хищников; у других подвижна губа, опрокидывающая опылителя на пыльник; третьи одурманивают насекомых специфическим запахом и т. п. Цветение орхидных продолжается, как правило, несколько месяцев .
Плод — сухая коробочка , раскрывающаяся 3 (6) створками, или ягода ; крайнее упрощение строения семян при значительном увеличении их числа. Коробочка содержит множество пылевидных семян, распространяющихся при помощи ветра. Двойного оплодотворения у орхидных нет, эндосперм не образуется, и поэтому для прорастания семян обязательно необходимо их заражение микоризным грибом.
Очень характерно формирование суккулентных , более или менее укороченных стеблей — так называемых псевдобульб, или туберидиев .
Гинецей орхидных состоит из трёх плодолистиков . Они образуют нижнюю завязь . У большинства видов она паракарпная , однокамерная, с париэтальной плацентацией . У наиболее примитивных орхидных — Apostasia , Neuwiedia ( Apostasioideae ) и некоторых более продвинутых таксонов из подсемейства Cypripedioideae — Phragmipedium , Selenipedium обнаружена синкарпная трёхгнёздная завязь с аксиальной плацентацией .
В основе андроцея орхидных лежит трёхчленный двухкруговой «лилейный» тип. Андроцей у ныне живущих орхидных претерпел кардинальные изменения по нескольким параметрам от этого исходного типа.
Число фертильных тычинок . Из шести фертильных тычинок андроцея лилейных лишь у наиболее примитивных орхидных осталось три: одна — из внешнего и две — из внутреннего кругов андроцея ( Neuwiedia , Apostasioideae ). Две фертильные тычинки внутреннего круга имеют виды подсемейства Cypripedioideae . У подавляющего большинства остальных видов семейства имеется лишь одна фертильная тычинка внутреннего круга.

Орхидеи имеют крошечные пылевидные семена . Их размеры колеблются в пределах 0,35—3,30 мм в длину и 0,08—0,30 мм в ширину. Ещё мельче зародыш — 0,05—0,26 мм в длину и 0,04—0,19 мм в ширину.
Эндосперм у орхидных претерпел сильные редукционные изменения. Он полностью отсутствует в зрелых семенах, вероятно, у всех представителей этого семейства. Некоторые зачатки эндосперма можно обнаружить лишь на ранних стадиях эмбриогенеза. Только у одной трети из эмбриологически изученных видов наблюдается образование многоклеточного (до 2—10 ядер) эндосперма. У многих видов образуется только первичное ядро эндосперма, которое дегенерирует без дальнейшего деления и абсорбируется развивающимся зародышем.
Орхидеи являются бесспорными лидерами среди цветковых растений по уровню семенной продуктивности. В одной коробочке у них может образовываться до четырёх миллионов семян.

Из жизненных форм доминируют многолетние травы, реже встречаются небольшие кустарники и крупные одревесневающие лианы . Наряду с автотрофными нередко встречаются сапрофитные формы .
Среди тропических видов в оранжерейных условиях выращивания нередко наблюдаются «долгожители» по 70 и более лет. Размеры у орхидных колеблются от нескольких миллиметров до 35 метров ( Vanilla planifolia ).
По характеру субстрата произрастания различают эпифиты , литофиты и наземные орхидные. Большинство орхидей — эпифиты. Литофиты и наземные жизненные формы составляют значительно меньшую группу. Они распространены преимущественно в умеренных широтах.
Все орхидные вступают в симбиоз с микоризообразующими грибами, которые поставляют растению воду и минеральные соли, получая взамен органические вещества. Есть также микотрофные орхидные, лишённые хлорофилла и фактически паразитирующие на грибе-симбионте (например, гнездовка , а также австралийская орхидея криптантемис , цветущая под землёй). Эпифитные орхидные, как правило, развивают многочисленные воздушные корни , лишённые волосков, зато снабжённые специализированной всасывающей тканью — веламеном . У некоторых эпифитных орхидных листья редуцируются, а функцию фотосинтеза принимают на себя уплощённые зелёные корни ( фаленопсис ) .
Зародыш в зрелых семенах орхидных полностью лишён эндосперма. Клетки зрелого зародыша содержат только белки и жироподобные вещества. Крахмальные зёрна полностью исчезают из клеток зародыша ко времени его созревания (Андронова, 1988). Таким образом, в зрелых семенах орхидных отсутствуют запасы углеводов, обеспечивающих энергетику делящихся клеток. Это обусловливает неспособность семян у подавляющего большинства видов орхидных к самостоятельному прорастанию без притока органических веществ извне.
Его прорастание и дальнейшее постсеменное развитие полностью обеспечивается за счёт определённых микоризообразующих грибов, которые проникают в семена. Иногда, если гриб окажется «сильнее» семечка орхидеи, он убивает его, но чаще орхидея и гриб живут в симбиозе. Некоторые орхидеи так приспособились к грибам, что могут жить без света под землёй, например гнездовка .

Система опыления у орхидных отличается высоким разнообразием как по агентам опыления (опылителям), так и по способам привлечения (аттрактации) последних на цветки. Основную роль в опылении орхидных играют насекомые. Главная отличительная особенность системы опыления орхидных — агрегация всей пыльцы пыльника в компактные массы поллинии. Для надёжного прикрепления поллиниев к телу опылителей и успешного их переноса на рыльце у орхидных выработались разнообразные механизмы. Основные их типы были детально изучены и изложены Ч. Дарвином (1884) в его замечательном труде «Различные приспособления, при помощи которых орхидеи опыляются насекомыми».
Особый интерес для исследователей системы опыления у орхидных представляют способы привлечения (аттрактации) опылителей на цветки. Пыльца, собранная в поллиниях, не может быть использована насекомыми для выкармливания потомства. К тому же у подавляющего большинства видов орхидных в нектаро-собирающих структурах (шпорцах) отсутствует нектар. Поэтому на безнектарные цветки орхидных насекомые-опылители привлекаются различными способами обманной аттрактации. У орхидных принято выделять следующие основные синдромы обманной аттрактации: сексуальное привлечение (pseudocopulation), цветковая мимикрия (flower mimicry), обман неопытных опылителей . Это разделение весьма условно, так как у многих видов орхидных нередко бывают задействованы сразу несколько способов обмана.
Сексуальный синдром привлечения опылителей в настоящее время известен у ряда родов орхидных Евразии ( Ophrys ), Австралии ( , , Cryptostylis , и др.) и Центральной Америки. Наиболее детально он изучен у видов рода Ophrys .
Цветковая мимикрия более широко распространена среди безнектарных орхидных. У орхидных с этим синдромом привлечения опылителей цветки или их отдельные части имитируют по окраске/форме/текстуре поверхности нектароносные цветки других растений или другие пищевые субстраты, используемые потенциальными опылителями.
Этот синдром в явной форме впервые был описан L. A. Nilsson (1980) у Dactylorhiza sambucina . Его характерными особенностями являются: яркий поисковый облик цветков/соцветий, наличие на губе ярких ложных указателей нектара (в виде контрастного рисунка или/и окрашенных папиллярных выростов), глубокий хорошо развитый шпорец.
После первых эмбриологических исследований орхидных была выдвинута гипотеза отсутствия у них двойного оплодотворения . Распространению этой гипотезы способствовал тот факт, что в зрелых семенах орхидных полностью отсутствует эндосперм. У орхидных неоднократно отмечены случаи невхождения второго спермия в центральную клетку зародышевого мешка. Нередко второй спермий входит в контакт с полярными ядрами и образует вместе с ними в центральной или базальной части зародышевого мешка ядерный комплекс (Навашин,1951; Савина, 1972). Однако примерно у одной трети видов орхидных наблюдается слияние второго спермия с центральной клеткой зародышевого мешка и образование первичной клетки эндосперма.
В коллекции орхидных Королевского ботанического сада Кью содержатся 24 образца орхидей, сохранившихся с начала XX века (их возраст около 100 лет); возраст растений 130 видов превышает 50 лет, а возраст растений 500 видов превышает, по крайней мере, 20 лет .
Многие виды одного рода и даже представители различных родов легко скрещиваются между собой, образуя многочисленные гибриды, способные к дальнейшему размножению. Большинство гибридов , появившихся за последние 100 лет, создано искусственно с помощью целенаправленной селекционной работы .
Первое искусственное межвидовое опыление произвёл . В 1856 году была создана первая гибридная орхидея Calanthe dominii ( × Calanthe furcata ). Джон Домини работал в течение двадцати лет и произвёл около 25 гибридов, в том числе, в 1863 году, первый межродовый гибрид exoniensis ( × ) .
На 2000 год существовало около 250 000 искусственных гибридов орхидей .
Правила и рекомендации по номенклатуре и регистрации гибридов и сортов орхидей основаны на положениях Международного кодекса ботанической номенклатуры и опубликованы в справочнике Номенклатура и регистрация орхидей .
Название гибрида состоит из трёх частей — родового названия ( рода или гибридного рода), названия грекса ( англ. grex ) и названия культивара ( клона ) (пишется в кавычках). Например: Paphiopedilum maudiae «Queen» ( Paphiopedilum lawrenceanum var. hyeanum × Paphiopedilum callosum var. sanderae ).
Клоном в цветоводстве называется конкретный представитель грекса, а также его вегетативное потомство (в том числе полученное при помощи массового меристемного клонирования ). Официальной международной регистрации подлежат именно грексы, в то время как клонам присуждаются различные награды многими национальными орхидными обществами. На клоны орхидей распространяются международные соглашения о сортах растений, в частности, преследуется фальсификация при продаже .
Названия межродовых гибридов создают из названий родов — партнёров по скрещиванию: Odontonia = Odontoglossum × Miltonia . Гибриды, созданные с использованием большого количества родов, иногда называют по имени селекционера: Vuylstekeara = Cochlioda × Miltonia × Odontoglossum (гибрид, зарегистрированный в 1911 году, назван в честь бельгийского садовода-селекционера C. Vuylsteke).
Аббревиатуры гибридных родов, используемых в промышленном и любительском цветоводстве , публикуются на сайте Королевского садоводческого общества . Последние изменения отражены в документе .
Одним из направлений селекционной работы является получение полиплоидных гибридов, для которых характерен относительно большой размер цветков. Наибольшее число полиплоидных гибридов зарегистрировано среди каттлей , ванд , фаленопсисов , цимбидиумов и у Dendrobium bigibbum .

Орхидеи известны в первую очередь как красивоцветущие декоративные растения , являются излюбленными оранжерейными растениями. Особую популярность снискали виды родов Каттлея , Дендробиум , Фаленопсис .
Плоды лианы ванили плосколистной ( Vanilla planifolia ) используются как ценная пряность («палочки ванили»).
Мясистые клубнекорни ряда орхидей (например, пальчатокоренника крапчатого ( Dactylorhiza maculata ), любки двулистной ( Platanthera bifolia ), кокушника длиннорогого ( ), анакамптиса пирамидального ( Anacamptis pyramidalis ) и других) содержат крахмал и применяются в медицине под наименованием клубней салепа (или салепа) ( лат. Tuber Salep ) как обволакивающее средство и для питания ослабленных больных. В зависимости от морфологических особенностей клубнекорней различают круглый салеп и пальчатый салеп .






